

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
2010年广州市甲型H1N1流感病毒分离株NA基因变异分析
引用本文
许沙沙, 常彦敏, 徐霖, 冯发深, 何霞, 王铸, 张定梅, 曹开源. 2010年广州市甲型H1N1流感病毒分离株NA基因变异分析. 检验医学, 2014, 29(1): 42-49[XU Shasha, CHANG Yanmin, XU Lin, FENG Fashen, HE Xia, WANG Zhu, ZHANG Dingmei, CAO Kaiyuan. Analysis on the NA gene variation of influenza A (H1N1) virus from Guangzhou in 2010. Labratory Medicine, 2014, 29(1): 42-49]
Permissions
2010年广州市甲型H1N1流感病毒分离株NA基因变异分析
摘要
目的 比较2010年从广州市分离到的甲型H1N1流感病毒神经氨酸酶(NA)基因与2009年中国大陆甲型H1N1流感病毒NA基因的变异情况, 为甲型H1N1流感的监测和防控提供参考资料。方法 收集2010年广州市有发热和呼吸道症状患者的咽拭子标本, 用甲型H1N1流感病毒特异性引物进行聚合酶链反应 (PCR) 检测, 扩增分离到的甲型H1N1流感病毒NA基因片段, 测序后与2009年的H1N1毒株进行比对和进化分析, 并用生物信息学方法对耐药位点和糖基化位点进行分析。结果 共收集1 194份咽拭子标本, 检测到甲型流感病毒阳性327份, 其中H1N1流感病毒6株, 与2009年分离的甲型H1N1流感病毒相比, 有16个位点发生了有义突变, 3个位点和NA活性相关, 其中222位氨基酸的变异位于NA活性位点上。结论 成功扩增了2010年广州市6株甲型H1N1流感病毒株NA基因并测序, 未发现H275Y耐药位点的变异。3毒株在NA活性位点222位、228位和425位等氨基酸位点处发生了变异, 需继续加强监测。
关键词:
甲型H1N1流感病毒; 神经氨酸酶; 耐药位点
Analysis on the NA gene variation of influenza A (H1N1) virus from Guangzhou in 2010
Abstract
Objective To compare the variations of neuraminidase (NA) gene of influenza A (H1N1) virus from Guangzhou in 2010 andNA gene of influenza A (H1N1) virus from Chinese mainland in 2009, and to provide the reference for the surveillance and prevention of influenza A (H1N1) virus.Methods Specimens were collected from patients with febrile respiratory tract symptoms from Guangzhou in 2010. The specimens were determined with influenza A (H1N1) virus specificity primers by polymerase chain reaction (PCR). The fragment ofNA gene was amplified. After sequencing, the results were compared with influenza A (H1N1) virus in 2009.The drug resistance sites and glycosylation sites were analyzed by biometric software.Results A total of 1 194 specimens were collected, and 327 influenza A virus strains were detected, including 6 strains were identified with influenza A (H1N1) virus. Compared with influenza A (H1N1) virus in 2009, there were 16 amino acid sites with variation. There were 3 amino acid sites relating with NA activity. However, the amino acid site 222 located in the NA activity sites.ConclusionsNA gene of the 6 strains of influenza A (H1N1) virus are amplified and sequenced successfully, and H275Y drug resistant site is not found. The amino acid site 222, 228 and 425 locating in NA activity sites are found with mutations in 3 strains. Therefore, the monitoring should be strengthened.
Keyword:
Influenza A ( H1N1) virus; Neuraminidase; Drug resistance site
流感病毒是有包膜的单股负链RNA病毒, 属于正黏病毒科 (orthomyxoviridae),根据核衣壳蛋白和基质蛋白不同, 分为甲 (A) 、乙 (B) 、丙 (C) 3型[ 1]。甲型H1N1流感于2009 年3月在墨西哥爆发, 最初被称为“人感染猪流感”, 世界卫生组织将流感大流行警告级别提高为6级[ 2],2009年4月中国卫生部发布公告, 明确将猪流感更名为“甲型H1N1流感”。 据世界卫生组织统计, 截至2010年3月19日, 这种新病毒已经波及213个国家, 共造成16 813例死亡[ 3]。许多学者认为华南地区是世界流感流行的一个主要起源地, 因此加强对甲型H1N1流感的监测对我国传染病防控具有重要意义[ 4]。神经氨酸酶 (neuraminidase, NA) 是流感病毒表面重要的糖蛋白, 是甲型流感病毒主要抗原之一, 对于流感病毒从感染细胞中的释放非常重要。为了监测甲型H1N1流感NA基因的变异情况, 及时发现具有流行病学意义的耐药株, 我们对2010年从广州采集的1 194份呼吸道标本中分离到的6株甲型H1N1病毒株NA基因进行测序, 与2009年的代表株进行比对, 对其耐药位点和同源性进行分析, 了解其变异情况, 为今后流感的监控和防治提供参考资料。
材料和方法
一、标本来源
2010年2到12月广州市中山大学附属二院和三院发热门诊病例咽拭子标本1 194份, 病例纳入标准为:发热3 d以内, 体温≥38 ℃;伴有咳嗽或咽喉疼痛等急性上呼吸道感染症状。
二、毒株
注:*为2009年世界卫生组织推荐的甲型H1N1流感病毒疫苗株
2010年本实验室分离的甲型H1N1流感病毒和2009年3月本实验室从广州市第1例甲型H1N1流感病毒感染患者体内分离的毒株, 名称为A/Guangdong/03/2009 (H1N1)(GenBank登录号GQ250161);从 GenBank中选取北京、上海、广东三地在2009年甲型H1N1流感流行时的NA序列共9株, 用以基因分型和比较, 见表1:
| 表1 NA基因用以进化分析的毒株 |
三、RNA的提取和cDNA的合成
对采集的咽拭子标本进行处理, 各取200 μL提取RNA (QIAmp MiniElute Virus Spin试剂盒),逆转录合成cDNA (Invitrogen, Superscript Ⅲ first strand逆转录试剂盒),操作步骤参照说明书进行, 反应条件为:25 ℃ 10 min, 50 ℃ 50 min, 85 ℃ 5 min, 37 ℃ 20 min。
四、甲型流感检测
对1 194份标本的cDNA全部进行甲型流感病毒检测。
1.引物 参照国家流感中心下发的引物, 目标片段为M基因, 上游引物5′-TTCTAACCGAGGTCGAAACG-3′, 下游引物5′- ACAAAGCGTCTACGCTGCAG-3′, 特异产物长度235 bp。
2.聚合酶链反应 (polymerase chain reaction, PCR) 反应体系 TaKaRa ExTaq mix 12.5 μL, 焦碳酸二乙酯水 7.5 μL, 上、下游引物各2 μL;cDNA模板1 μL。 反应条件:95 ℃ 5 min, 94 ℃ 30 s, 52 ℃ 30 s, 72 ℃ 1 min, 35个循环, 72 ℃ 10 min。
五、甲型H1N1流感病毒检测
对甲型流感病毒检测阳性的标本进行甲型H1N1流感病毒特异PCR检测。
1. 引物 参照国家流感中心下发的引物, 目标片段为HA基因, 上游引物5′-AATAACATTAGAAGCAACTGG-3′, 下游引物5′-AGGCTGGTGTTTATRGCACC-3′, 特异产物长度153 bp。
2.PCR 同上。
六、NA克隆和序列测定
参考GenBank中甲型H1N1流感病毒株NA序列 (GenBank 登陆号GQ250162) 分别设计分段引物NA-1和NA-2, NA-1引物扩增1~643 bp, NA-2引物扩增505~1 410 bp,NA基因全长1 410 bp, 引物序列见表2:
| 表2 NA基因扩增引物 |
PCR反应体系:TaKaRa ExTaq mix 25 μL, 焦碳酸二乙酯水 15 μL, 上下游引物各4 μL, cDNA模板2 μL。 反应条件:95 ℃ 5 min, 94 ℃ 30 s, 55 ℃ 30 s, 72 ℃ 1 min, 35 个循环, 72 ℃ 10 min。
胶回收试剂盒 (TAKATA) 回收PCR产物, 连接到pMD 18-T Vector (TAKATA),转化DH-5α感受态细胞, 用Amp+LB筛选阳性菌落提取重组质粒, PCR初步鉴定重组质粒, 送上海英骏公司测序。
七、变异分析
利用 Clustal 1.83软件[ 5]对测序毒株HA基因序列和氨基酸序列进行比对和分析, 利用Glyc 1.0 Server (http://www.cbs.dtu.dk/services/NetNGlyc/)在线软件预测其糖基化位点, 并对其耐药位点进行分析。
八、系统进化分析
将分离的6株广州甲型H1N1流感病毒分离株和从GenBank中筛选的9株甲型H1N1流感病毒的HA基因用MEGA4.0软件[ 6]近邻连接法 (neighbor-joining method) 构建系统进化树。
结 果
一、标本流行病学资料
1 194份咽试子标本中检测甲型流感病毒327份, 其中检测到H1N1流感病毒6株。
四、甲型H1N1流感病毒的NA 基因PCR扩增及测序
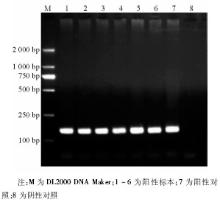
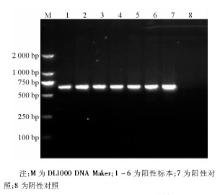
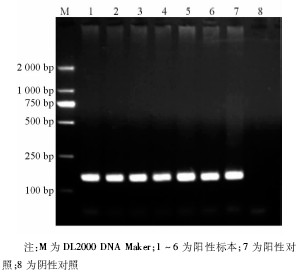
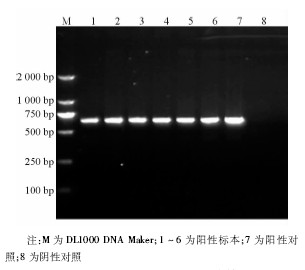
6株甲型H1N1流感病毒分离株NA-1段基因的逆转录PCR产物全长643 bp, 见图3。6株甲型H1N1流感病毒的NA-2 段基因的逆转录PCR产物全长905 bp, 与理论值相符, 见图4。回收PCR扩增产物, 与T载体连接, 由上海英骏公司进行测序 (结果略) 。
 | 图3 NA-1片段逆转录PCR电泳结果 |
五、变异分析
注:ID为同一株病毒比较的空白值
注:*为本实验室2010年分离甲型H1N1流感毒株;**2009年世界卫生组织规定的疫苗株.
1.NA基因同源性分析 2009年流感病毒代表株和2010年流感病毒分离株共15株, 将其NA基因共1 410个碱基, 用Clustalx 1.83进行序列比对, 用BioEdit软件得到15株NA基因的氨基酸相似性矩阵, 见表3。2010年分离株和2009年代表株氨基酸相似性较高, 介于0.985~0.999。
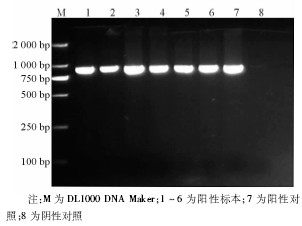 | 图4 NA-2片段逆转录PCR电泳结果 |
| 表3 NA核酸相似性矩阵 |
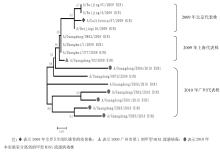
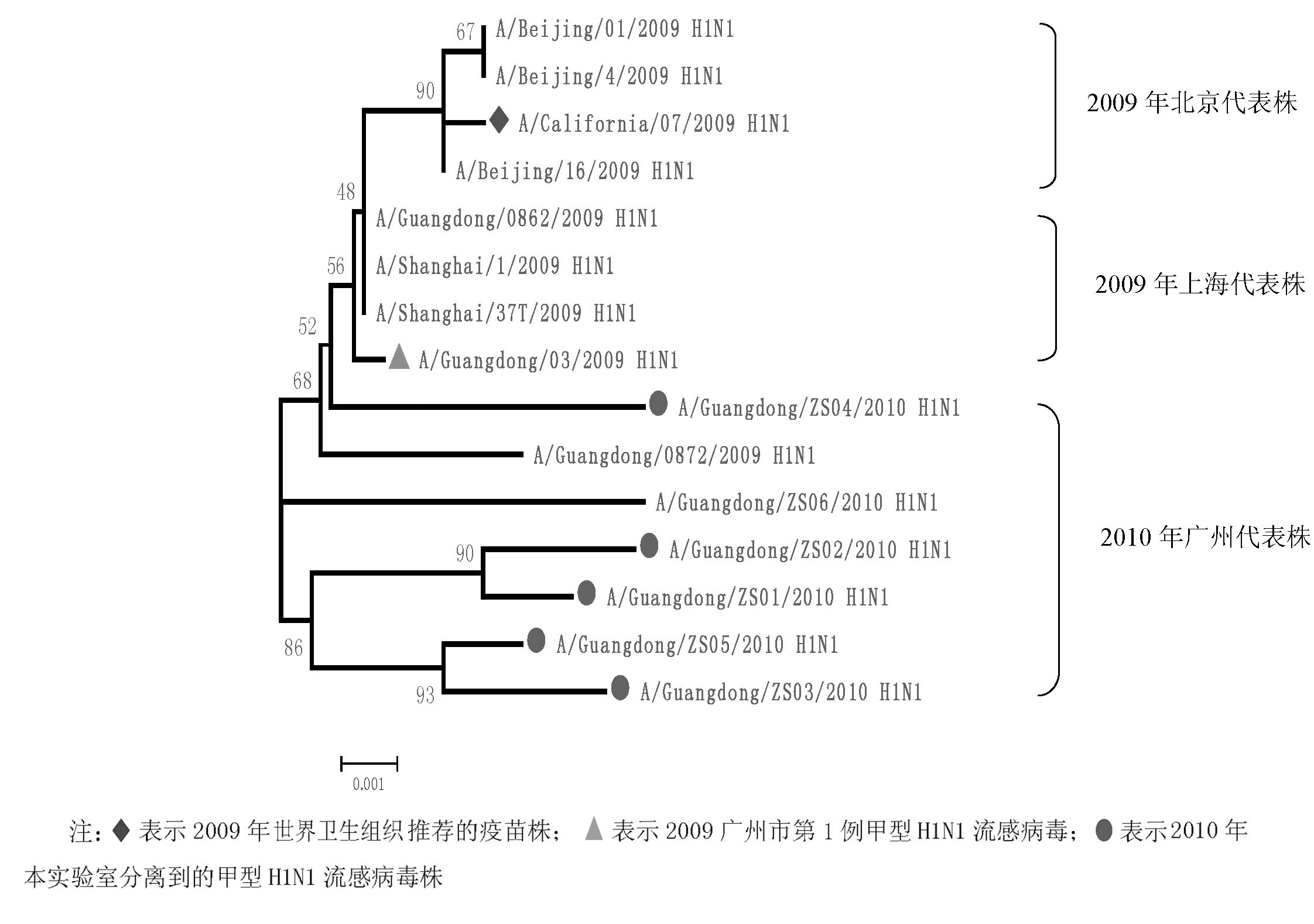
2.NA系统进化树分析 根据NA基因全长序列 (1 410 bp),对2009年代表株和2010年分离株共15株甲型H1N1流感病毒株进行了系统进化分析, 包括本研究分离到的6株, 还有选取的2009年代表株。在系统进化树上, 15株毒株分成了3个分支, 见图5:
 | 图5 甲型H1N1流感病毒NA基因系统进化树 |
3.氨基酸变异分析 2010年分离的6株甲型 H1N1 流感病毒的NA基因与2009年中国大陆的代表株比对, 6株分离株共有30个碱基位点发生突变, 其中有义突变为16个, 3个位点和NA活性相关, 其中222位氨基酸的变异位于NA活性位点上, 未发现H275Y耐药位点的变异, 见表4:
| 表4 2010年分离株和2009年代表株NA片段氨基酸比对结果 |
4.糖基化位点变异分析 分别将2010年广州市分离到的6株甲型H1N1病毒株和2009年代表株NA基因进行糖基化位点的预测。结果发现4株甲型H1N1病毒株的糖基化位点和2009年代表株相比并没有明显的变化。
5.NA蛋白结构建模 通过蛋白分析专家系统EXPASY (http://ca.expasy.org/)提供的蛋白组学和序列分析工具SWISS-MODEL, 模拟构建各NA蛋白的空间构象。3株病毒株在NA活性位点222位、228位和425位等氨基酸位点处发生了变异, 见图6:
 | 图6 NA蛋白三维结构 |
讨 论
NA蛋白是流感病毒包膜上重要的糖蛋白, 能切断细胞表面的唾液酸, 促使病毒从感染细胞膜上释放, 防止子代病毒自身凝集, 促进病毒扩散并增强其感染能力。由于NA在流感病毒复制和传播中起重要作用, 且其活性中心的氨基酸组成高度保守, 因此是研制抗流感药物的重要靶点。
目前抗流感药物有2种:一种为M2离子通道阻滞剂, 如金刚烷胺, 但是其副作用很大, 而且很容易产生耐药株, 有学者报道现在流感病毒对烷胺类的耐药现象十分严重[ 7, 8]。而2009年甲型H1N1流感病毒对烷胺类耐药情况同样严重, 主要是在NA第31位氨基酸的变异 (S31N)[ 9]。在这种情况下NA抑制剂成为目前抗流感病毒研究的热点[ 10],而且奥司他韦对甲型流感和乙型流感都有效, 不易引起耐药性且耐受性好。但是自从奥司他韦应用于临床以来, 不断有耐药株出现, 耐药率逐年上升, 耐药率在成人为0.4%~1%[ 11],在儿童为4%~8%,其中日本儿童的耐药率高达18%。
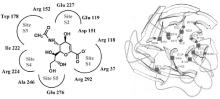
流感病毒NA的活性位点结构示意图见图7[ 12, 13],NA催化位点以及周围相关位点高度保守[ 14, 15]。直接参与催化作用的活性区域有5个, 分别为 S1~S5, S1由3个氨基酸残基 (R118、R292、R371) 组成, S2由2个氨基酸残基 (E227、E119) 组成, S3由2个氨基酸残基 (W178、I222) 组成, S4由3个氨基酸残基 (I222、R224、A246) 组成, S5由2个氨基酸残基 (A246、E276) 组成。间接参与催化作用的位点有R156、Y406、S179、D198、E228、N294、E425等氨基酸位点。
 | 图7 流感病毒NA的活性位点结构示意图 |
这些位点都直接或间接地参与NA的催化, 所以此处氨基酸如果发生突变, 就会对NA的活性产生影响, 减低流感病毒对NA抑制剂的敏感性, 甚至会导致对NA抑制剂的耐药。通过对本研究分离到的6株甲型H1N1流感病毒的NA基因的测序, 氨基酸序列分析发现部分毒株出现N42S、N59D、Q313K等的氨基酸位点变异, 这些均未涉及NA催化位点。但是在A/Guangdong/ZS03/2010 (H1N1) 毒株中出现E228G氨基酸突变, A/Guangdong/ZS04/2010 (H1N1) 毒株出现E425G氨基酸突变, 这些位点都与NA的活性有关, 但这些毒株其耐药性是否有变化, 有待进一步研究。
同时在A/Guangdong/ZS01/2010 (H1N1) 毒株中出现N222S氨基酸的突变 (虽然与文献报道I222位突变的碱基不一样, 但是222位氨基酸还是存在突变的) 。许多研究表明222位氨基酸的改变会导致流感病毒对NA抑制剂的耐药[ 16]。因此需继续对其耐药性密切监测。
从NA三维结构图中可以看到NA活性位点组成一个疏水性的口袋状结构, 可以结合NA抑制剂奥司他韦等药物, 起到抗病毒的作用。如果疏水性口袋状结构的氨基酸位点发生变异, 会导致无法结合奥司他韦等药物, 则会产生流感病毒耐药株。尽管结构图中只有少部分氨基酸的突变, 但突变位点在NA活性位点上可能对其耐药性产生影响, 需后续试验继续研究。
许多研究表明, 275位氨基酸的变异会对奥司他韦产生耐药[ 17],通过比较分析2010年分离到的甲型H1N1流感病毒, 均未发现275位的变异, 提示在2010年广州散发的甲型H1N1流感病毒对NA抑制剂仍然敏感。2010年分离株NA糖基化位点与2009年代表株比较并没有变化, 与Saxena等[ 18]的报道相同。同时发现2010年分离株与2009年代表株同源性较高 (0.985~0.999),提示NA基因仍是同一来源, 未发生较大变异。对NA系统进化树分析发现, 2009年代表株为一支, 而2010年分离株为一支, 说明2010年与2009年相比,NA基因有部分碱基发生变异。
通过以上分析, NA蛋白275位氨基酸并未出现变异, 提示还没有出现对奥司他韦的耐药株, 但是在NA活性中心周围已经存在有变异的氨基酸, 可能会对其耐药性有影响。同时随着奥司他韦在临床上的广泛应用, 甲型H1N1流感病毒耐药株在世界上其他国家不断有报道, 更应该继续加强对甲型 H1N1流感病毒的耐药性监测以及序列分析工作, 在分子遗传进化分析的基础上, 及时发现有流行病学意义的耐药株, 为国家制定甲型 H1N1 流感防控策略提供参考。
参考文献
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|