
{kind=link}
HBV感染者外周血Th17细胞和Treg细胞的检测及意义
引用本文
胡月明, 马莉, 李虹. HBV感染者外周血Th17细胞和Treg细胞的检测及意义. 检验医学, 2014, 29(1): 21-25[HU Yueming, MA Li, LI Hong.. The detections of peripheral blood Th17 cells and Treg cells in HBV infected patients and their significance. Labratory Medicine, 2014, 29(1): 21-25] 
Permissions
HBV感染者外周血Th17细胞和Treg细胞的检测及意义
摘要
目的 探讨乙型肝炎病毒 (HBV) 感染者外周血中辅助性T细胞17 (Th17) 细胞和调节性T细胞 (Treg) 的比例变化及其与HBV感染后肝脏损伤的关系。方法 采用流式细胞术检测34例乙型肝炎病毒携带者 (ASC) 、50例慢性乙型肝炎 (CHB) 患者、32例肝细胞癌 (HCC) 伴活动性肝炎 (简称HCC) 患者及30名健康对照者 (正常对照组) 外周全血Th17细胞和Treg细胞, 同时采用西门子1650全自动生化分析仪检测血清丙氨酸氨基转移酶 (ALT) 、天门冬氨酸氨基转移酶 (AST) 。结果 与正常对照组的Th17细胞比例 (1.13%±0.37%)比较, ASC组略有升高 (1.30%±0.28%),但差异无统计学意义(P>0.05);CHB组 (2.06%±1.15%)和HCC组 (5.89%±3.31%)均明显升高(P<0.01),且HCC组高于CHB组(P<0.05) 。Th17细胞比例与ALT、AST呈正相关[相关系数(r)分别为0.682、0.637,P均<0.01]。与正常对照组CD4+CD25+CD127lowTreg细胞 (7.18%±2.28%)比较, ASC组 (7.72%±2.05%)略有升高, 但差异无统计学意义(P>0.05);CHB组 (9.01%±2.92%)和HCC组 (10.86%±1.77%)均明显升高(P<0.01),且HCC组高于CHB组(P<0.05) 。Treg细胞与ALT、AST均呈正相关[r值分别为0.452(P<0.01) 、0.307(P<0.05)]。外周血Th17细胞比例与Treg细胞比例呈正相关(r=0.327,P<0.01) 。正常对照组Th17/Treg最低, 为0.17±0.16, ASC组略有上升 (0.18±0.13),但差异无统计学意义(P>0.05);CHB组 (0.23±0.19) 和HCC组 (0.52±0.54) 明显升高(P<0.05、P<0.01) 。结论 HBV感染后随着疾病的发生、发展, 外周血Th17细胞和Treg细胞的比例增加。由于Th17细胞和Treg细胞构成的平衡被打破, 趋向Th17细胞所介导的炎症反应, 引起肝细胞损伤。
关键词:
辅助性T细胞17; 调节性T细胞; 乙型肝炎病毒
The detections of peripheral blood Th17 cells and Treg cells in HBV infected patients and their significance
Abstract
Objective To investigate the percentage changes of T helper 17 (Th17) cells and regulatory T (Treg) cells in the peripheral blood of hepatitis B virus (HBV) infected patients and their correlations with liver injury after HBV infection.Methods The peripheral blood of 34 asymptomatic HBV carrier (ASC), 50 chronic hepatitis B (CHB) patients and 32 hepatocellular carcinoma (HCC) patients were collected. A total of 30 healthy controls (control group) were enrolled. The percentages of Th17 and Treg cells were detected by flow cytometry. Alanine aminotransferase (ALT) and aspartate aminotransferase (AST) were detected by Bayer ADVIA 1650 automatic biochemistry analyzer.Results Compared with control group (1.13%±0.37%), the percentage of Th17 cells in the peripheral blood of ASC group (1.30%±0.28%) was higher with no statistical significance (P> 0.05), but it obviously increased in CHB group (2.06%±1.15%) and HCC group (5.89%±3.31%,P<0.01). The percentage of Th17 cells in HCC group was higher than that in CHB group (P<0.05). The percentage of Th17 cells was positively correlated with ALT and AST [coefficients of correlation(r) were 0.682 and 0.637,P<0.01]. Compared with control group (7.18%±2.28%), the percentage of CD4+CD25+CD127lowTreg cells in the peripheral blood of ASC group (7.72%±2.05%) was higher with statistical significance (P> 0.05), but it increased obviously in CHB group (9.01%±2.92%) and HCC group (10.86%±1.77%,P<0.01), and that in HCC group was higher than that in CHB group(P<0.05). The correlation of Treg cells was positive with ALT and AST (r=0.452,P<0.01;r=0.307,P<0.05). The correlation of Th17 cell percentage was positive with Treg cell percentage (r=0.327,P<0.01). The rate of Th17/Treg in control group was the lowest (0.17±0.16), that in ASC group (0.18±0.13) increased slightly, and there was no statistical significance between control group and ASC group (P> 0.05). However, there was obviously increasing in CHB group (0.23±0.19,P<0.05) and HCC group (0.52±0.54,P<0.01).Conclusions In HBV infected patients, the percentages of Th17 and Treg cells increase. As for the immune balance of Th17 and Treg cells is broken, the increasing of Th17 cells mediates immune injury and induces the inflammation of liver cells.
Keyword:
T helper 17 cell; Regulatory T cell; Hepatitis B virus
乙型肝炎是全球广泛流行的传染性疾病。大量研究表明, 乙型肝炎的发病机制不是单独由乙型肝炎病毒 (hepatitis B virus, HBV) 直接引起的肝细胞损伤, 而是HBV感染后引发机体细胞免疫应答和病毒共同作用的结果。因此, 由细胞免疫介导的免疫损伤成为HBV感染肝细胞损伤机理研究的热点。
辅助性T细胞亚群在免疫系统中扮演很重要的角色, 可产生多种细胞因子, 参与机体抗病毒的免疫应答, 对病毒清除起重要作用。辅助性T细胞17 (T help 17 cells, Th17) 是不同于Thl和Th2的一种CD4+T细胞亚型, 因其产生大量白细胞介素-17 (IL-17) 而命名。最新研究表明Th17细胞可以调节和控制免疫应答[ 1],其主要作用于感染组织, 发挥募集中性粒细胞的生物学功能, 促进炎性反应, 对宿主抵抗特殊病原体发挥重要作用。与Th17 细胞不同, 调节性T细胞 (regulatory T cells, Treg) 是一类具有免疫抑制作用的CD4+T细胞亚群, 能够控制免疫应答的强度, 减轻对机体组织的免疫损伤。Th17和Treg细胞分别起着促进与抑制炎症反应的作用。促炎性Th17细胞与抑制性Treg细胞之间平衡状态的打破是很多炎症及自身免疫性疾病发病的关键因素之一[ 2]。近年来, 临床对很多炎症性疾病, 例如炎性肠病、视网膜炎和心肌炎患者体内促炎性Th17细胞与抑制性Treg细胞间平衡作用的研究都已取得突破性进展[ 3, 4, 5]。对CHB患者体内Th17细胞或Treg细胞的研究也已展开。然而, 慢性HBV感染后不同临床类型患者体内Th17细胞和Treg细胞平衡作用的研究鲜有报道。
为探讨Th17细胞和Treg细胞介导的免疫反应与乙型肝炎发生发展的关系, 我们选取HBV感染后肝细胞不同程度损伤的患者, 检测其外周血Th17和Treg细胞比例的变化, 探讨其介导的免疫反应与乙型肝炎病理损伤的可能关联性, 以期为临床治疗的监测和预后的判断提供依据。
材料和方法
一、研究对象
收集2011年12月至2012年7月贵阳医学院附属医院感染科及贵阳市第五人民院肝病科门诊及住院患者116例, 所有患者的诊断均符合中华医学会肝病学分会、感染病学分会《慢性乙型肝炎防治指南2011年更新版》的诊断标准[ 6],同时排除合并其他类型肝炎病毒感染、酒精性肝病、自身免疫性肝炎等疾病, 无既往病史。
1. 无病状乙型肝炎病毒携带者 (asymptomatic hepatitis B virus carrier, ASC) 组 ASC患者34例, 男22例, 女12例, 年龄28~41岁。
2. 慢性乙型肝炎 (chronic hepatitis Bpatient, CHB) 组 CHB患者50例, 男38例, 女12例, 年龄14~72岁。
3. 肝细胞癌 (hepatocellular carcinoma, HCC) 伴活动性肝炎组 (简称HCC组) HCC患者32例, 男21例, 女11例, 年龄24~51岁。所选患者近期未接受干扰素及抗病毒药物治疗, 未进行放化疗。
4. 正常对照组 选取贵阳市第一人民医院健康体检者30名, 男16名, 女14名, 年龄15~56岁。入选标准:无糖尿病、结核病、自身免疫性疾病和近期感染史;无肝炎症状和体征, 血压正常, 心肺功能正常;乙型肝炎表面抗原 (HBsAg) 阴性, 抗丙型肝炎病毒 (HCV) 阴性。
二、标本采集
以无菌方式采集所有对象晨起空腹静脉血10 mL, 平均分为2份。一份不加抗凝剂, 分离血清后用于检测ALT、AST水平;另一份采用EDTA-K2抗凝, 用于细胞分离培养后流式细胞仪检测。所有标本采集前都征得研究对象的知情同意和各医院医学伦理委员会的批准。
三、主要试剂及仪器
1主要试剂 异硫氰酸荧光素 (FITC)-抗人CD3抗体、藻红蛋白花青苷7 (PECy7)-抗人CD8抗体、藻红蛋白 (PE)-抗人IL-17抗体、抗人CD69抗体、Treg Cocktail、PECy7-抗人IgG1抗体、PE-抗人IgG2a抗体、破膜剂 (perm2) 、红细胞裂解液、磷酸盐缓冲液 (PBS) 均购于美国BD Bioscience公司。RPMI 1640培养基 (Lonza Bio Whittaker);离子霉素 (ionomycin, IO) 、 (12-)十四酸佛波酯(-13-)乙酸盐 (phorbol 12-myristate-13-acetate, PMA) 、莫能霉素均购于美国Sigma公司。ALT和AST检测试剂盒购自德国Siemens公司。
2.仪器 FACSCanto Ⅱ流式细胞仪 (美国Becton-Dickson公司);西门子1650全自动生化分析仪 (德国Siemens公司) 。
四、方法
1.Th17细胞的检测 取新鲜无菌EDTA-K2抗凝全血和RPMI1640培养液各250 μL混匀, 分别加入2.5 μL PMA (终浓度50 ng/mL) 、5 μL离子霉素 (终浓度1 μg/mL) 和0.8 μL莫能霉素 (终浓度1.7 μg/mL),置于37 ℃、5%CO2刺激培养5 h。取培养后全血细胞100 μL分别加入荧光素标记单克隆抗体CD3-FITC 20 μL、CD8-PECy7 5 μL, 室温避光孵育30min。 加入红细胞裂解液裂解红细胞, PBS洗涤。加入固定破膜剂1 000 μL, 室温破膜后PBS洗涤。再加入20 μL抗人IL-17-PE抗体, 室温避光孵育, PBS洗涤后立即上流式细胞仪机检测。同时进行同型阴性对照处理。以前向角 (FSC) 对侧向角 (SSC) 圈出淋巴细胞群, 以CD3+和CD8+细胞设门, 从中圈出CD3+CD8-细胞, 选择CD3+CD8-IL-17+细胞群, 使用IL-17同型对照调节电压及补偿, 即得出Th17细胞占外周血CD4+淋巴细胞的比例。
2.Treg细胞的检测 取新鲜无菌EDTA-K2抗凝全血100 μL, 再加入Treg cooktail试剂20 μL, 室温避光孵育, 裂解红细胞, PBS洗涤。立即上流式细胞仪机检测。同时进行同型阴性对照处理。以FSC对SSC圈出淋巴细胞群, 从中圈出CD4+CD25+细胞群, 再圈出CD4+CD25+CD127low细胞, 以CD25为同型对照调节电压及补偿, 记录Treg细胞占外周血CD4+淋巴细胞的比例。
3.流式细胞术 (FCM) 分析 以BD FACSTM7-Color Setup Beads和BDTMCytomter Setup Tracking Beads调整流式细胞仪, 启动FACSCanto软件, 使仪器处于最佳工作状态。采用FACSDiva软件检测及分析。
4.血清丙氨酸氨基转移酶 (ALT) 、天门冬氨酸氨基转移酶 (AST) 水平检测 采用西门子1650全自动生化分析仪检测血清ALT、AST水平, 严格按照试剂操作说明书进行。
五、统计学方法
采用SPSS13.0统计软件进行分析, 计量资料以±s表示。组间采用多因素方差分析, 相关性比较采用Sperman相关性检验。检验水准χ2=0.05,P<0.05为差异有统计学意义。
结 果
二、HBV感染者外周血CD3+、CD3+CD4+ T细胞比例变化
注:与正常对照组比较,*P<0.05、**P<0.01;与ASC组比较,#P<0.05、##P<0.01
与正常对照组外周血CD3+、CD3+CD4+ T细胞比例比较, ASC组、CHB组、HCC组3组均明显降低(P<0.05、P<0.01) 。与ASC组患者外周血CD3+、CD3+CD4+ T细胞比例比较, CHB组及HCC组明显降低(P<0.05、P<0.01) 。见表2:
| 表2 HBV感染者和正常对照组外周血CD3+、 CD3+CD4+ T细胞检测结果(±s,%) |
注:与正常对照组比较,*P<0.05、**P<0.01;与ASC组比较,#P<0.05、##P<0.01
三、HBV感染者外周血Th17细胞和Treg细胞比例的变化
注:与正常对照组比较,*P<0.05、**P<0.01;与ASC组比较,#P<0.05、##P<0.01;与CHB组比较, ▲P<0.05
与正常对照组外周血Th17细胞比例比较, ASC组稍高, 但差异无统计学意义(P>0.05);CHB组、HCC组明显升高(P<0.01),且高于ASC组(P<0.05、P<0.01); HCC组患者外周血Th17细胞比例高于CHB组(P<0.05) 。外周血Treg细胞的比例变化趋势与Th17细胞相同, 见表3、图1。
HBV感染者外周血Th17细胞和Treg细胞呈正相关(r=0.327,P<0.01) 。正常对照组Th17/Treg比值最低, ASC组稍高, 但二者之间差异无统计学意义(P>0.05);CHB组和HCC组均明显高于正常对照组(P<0.05、P<0.01);HCC组明显高于ASC组和CHB组(P<0.05),见表3:
| 表3 各组外周血Th17细胞和Treg细胞比例检测结果比较(±s) |
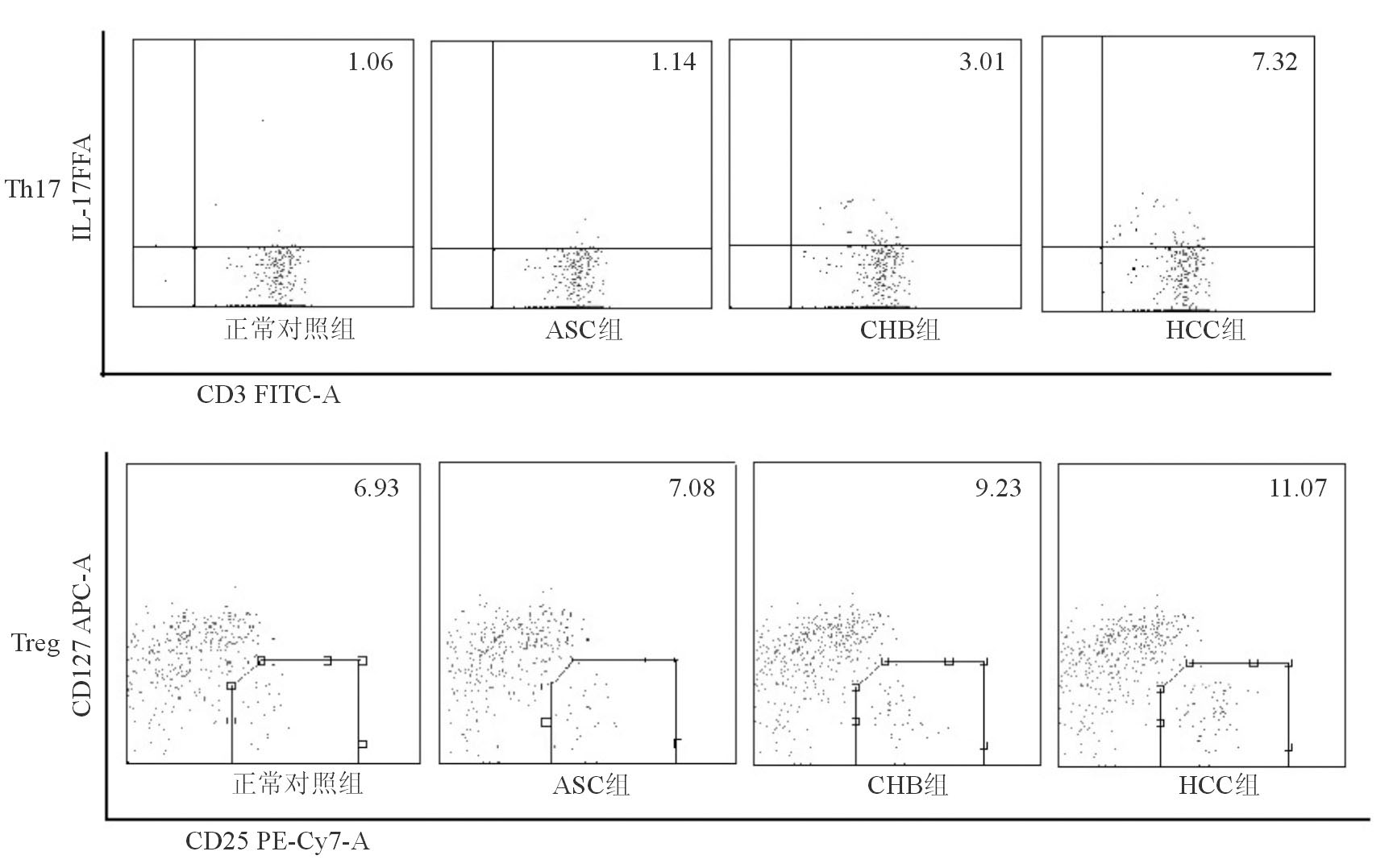 | 图1 HBV感染者及正常对照组外周血Th17细胞、Trge细胞比例 |
讨 论
细胞免疫在机体控制HBV感染是至关重要的, 其效应形式主要有2种:与靶细胞特异性结合, 破坏靶细胞膜, 直接杀伤靶细胞;另一种是释放细胞因子, 抵御感染、肿瘤等疾病。本研究对HBV感染患者外周血CD3+、CD3+CD4+ T细胞比例进行分析, 结果发现CHB和HCC患者外周血CD3+、CD3+CD4+ T细胞比例明显低于ASC组和正常对照组, 与文献报道[ 8]结果一致。其认为CD3+CD4+ T细胞生成减少。可以导致特异性抗体产生不足, 不能清除游离的HBV, 这可能是造成病毒在体内持续感染、患者病情加重的原因。
Th17细胞是最早参与抗感染免疫的效应性T细胞。研究表明, 机体适应性免疫系统可以通过IL-17和固有免疫系统相联系, 并促进炎症反应过程。为探究HBV感染后Th17细胞与乙型肝炎病情的关系, 本研究对不同HBV感染阶段的患者外周血Th17细胞比例变化, 并结合患者肝脏功能指标进行了研究。实验结果显示, 除ASC者外, CHB组和HCC组患者外周血Th17细胞比例均明显升高, HCC组Th17细胞比例较CHB组升高更为明显。这些均表明HBV感染后Th17细胞在T辅助细胞中所占的比例上升, 且外周血Th17细胞比例与血清ALT和AST水平呈正相关。提示HBV感染后患者体内Th17细胞参与肝细胞损伤, 引起肝脏炎症反应;并与HBV感染后肝细胞炎症加重有关。
1995 年日本学者发现, CD4+CD25+T细胞回输之后可以抑制裸鼠多种自身免疫性疾病的发生, 该细胞被命名为Treg细胞。Treg细胞能够下调机体免疫应答的水平, 发挥免疫抑制作用, 控制免疫应答的强度, 减轻对机体组织损伤以维持自身耐受。为探讨HBV感染后Treg细胞与肝脏功能的关系, 本研究也对各组实验对象外周血Treg细胞的比例变化进行了检测。结果显示ASC患者外周血Treg细胞比例与正常对照组比较差异无统计学意义, 而CHB和HCC患者外周血Treg细胞比例增高, 与Peng等[ 2]的报道一致。Stoop等[ 9]报道CD4+CD25+Treg细胞与HBV特异性T细胞的增殖反应有关, 去除Treg细胞后, HBV特异性T细胞增殖反应增强, 提示Treg细胞能抑制 HBV 特异性免疫反应。郑峻松等[ 10]研究发现, Treg细胞对效应性T细胞的作用通过接触机制直接作用, 进行特异性免疫抑制。那么HBV感染者外周血Treg细胞比例的增高对效应性T细胞——Th17细胞是否有抑制作用呢?
为深入研究这一问题, 本研究对受试者外周血Th17细胞和Treg细胞比例进行相关性分析, 发现Th17细胞和Treg细胞比例的变化呈正相关(r=0.327,P<0.01) 。究其原因可能为HBV感染导致患者体内效应性Th17细胞增多, 为拮抗效应性T细胞对机体的损伤, 抑制性Treg细胞反馈性增多。另外, Th17细胞和Treg细胞的分化发育过程中都受到TGF-β的调节。朱杰等[ 11]的研究结果表明CHB患者血清及肝组织中TGF-β均升高, 邰伯军等[ 12]也证实HCC患者体内TGF-β显著增高。因此, 本研究中CHB和HCC患者外周血中Th17细胞和Treg细胞的比例同时增高也可能是体内TGF-β升高, 促进其分化的结果。
Th17细胞和Treg细胞具有共同的起源, 相互构成复杂的作用, 形成一个网络。Th17细胞和Treg细胞之间平衡状态的打破可能是很多炎症性疾病及自身免疫性疾病发生和发展的一个关键因素。本研究发现在所有受试者中正常对照组Th17/Treg比值最低, 在HBV感染患者中随着肝细胞炎症的加重Th17/Treg比值逐渐升高。进一步表明尽管HBV感染后外周血中Th17细胞和Treg细胞的比例均同时升高, 但是其升高的幅度是不一致的;随着肝细胞炎症的加重, 外周血Th17细胞比例的升高更为明显, 导致Th17细胞和Treg细胞平衡度破坏。我们认为HBV感染患者外周血Th17细胞比例增高与肝细胞炎症损伤加重可能互为因果, 这也是患者病情进一步发展的因素之一。
综上所述, HBV感染后随着疾病的发生、发展, 外周血Th17细胞和Treg细胞比例明显增高。由于Th17细胞比例增高更为明显, 使其与Treg细胞构成的网络平衡被打破, 机体趋向于Th17细胞所介导的炎症反应, 造成肝细胞损害, 参与疾病的发生、发展。因此, 外周血Th17细胞和Treg细胞比例的检测可作为HBV感染后指导临床免疫治疗的指标。
参考文献
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|