


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
佛波酯诱导血小板黏附受体GPⅠbα酶切的机制研究
引用本文
王志成, 罗梅宏, 谢如锋, 张晓峰. 佛波酯诱导血小板黏附受体GPⅠbα酶切的机制研究. 检验医学, 2013, 28(5): 420-424
WANG Zhicheng, LUO Meihong, XIE Rufeng, ZHANG Xiaofeng. Research on molecular mechanisms of PMA-induced platelet GPⅠbα shedding. Labratory Medicine, 2013, 28(5): 420-424
Permissions
WANG Zhicheng, LUO Meihong, XIE Rufeng, ZHANG Xiaofeng. Research on molecular mechanisms of PMA-induced platelet GPⅠbα shedding. Labratory Medicine, 2013, 28(5): 420-424
Copyright©2013, 《检验医学》编辑部
《检验医学》编辑部版权所有
佛波酯诱导血小板黏附受体GPⅠbα酶切的机制研究
作者简介:王志成,男,1975年生,博士,助理研究员,主要从事血小板活化和凋亡以及血小板储存损伤机制研究。
摘要
目的 佛波酯(PMA)能诱导血小板黏附受体GPⅠbα酶切,但分子机制未完全阐明。本研究主要探讨PMA诱导GPⅠbα酶切的分子机制。方法 取健康志愿者静脉血,分离得到洗涤血小板。将洗涤血小板分别与蛋白激酶C(PKC)抑制剂、活性氧(ROS)抑制剂、NAD(P)H氧化酶抑制剂、线粒体ROS拮抗剂或二甲基亚矾(DMSO)对照等预孵育,再与PMA孵育。流式细胞仪检测ROS浓度;Western blot检测GPⅠbα酶切产物;用非同位素标记法检测PKC活性。结果 PKC抑制剂BIM和ROS抑制剂二硫苏糖醇(DTT)单独使用时,都能完全抑制PMA诱导的GPⅠbα酶切。PMA浓度依赖地诱导血小板ROS产生,PKC抑制剂和NAD(P)H氧化酶抑制剂都能抑制PMA诱导的ROS产生,而线粒体ROS拮抗剂没有抑制作用。BIM抑制PMA诱导的PKC活化,DTT则没有抑制作用。结论 PMA可能通过PMA-PKC-NAD(P)H氧化酶-ROS-ADAM17-GPⅠbα信号通路调节GPⅠbα酶切。
关键词:
血小板; GPⅠbα酶切; 佛波酯; 蛋白激酶C; 活性氧
中图分类号:R446.11
文献标志码:A
文章编号:1673-8640(2013)05-0420-05
Research on molecular mechanisms of PMA-induced platelet GPⅠbα shedding
Abstract
Objective Phorbol 12-myristate-13-acetate (PMA) could induce platelet GPⅠbα shedding, however, the molecular mechanisms are not fully elucidated. This study investigates the molecular mechanisms of PMA-induced platelet GPⅠbα shedding. Methods Washed platelets were obtained from anti-coagulated healthy volunteers' whole blood by centrifugation and washing. Washed platelets were pre-incubated with protein kinase C (PKC) inhibitor, reactive oxygen species (ROS) antagonist, NAD(P)H oxidase inhibitor, mitochondrial ROS antagonist, or dimethyl sulfoxide (DMSO), and were incubated with PMA. ROS levels were measured by flow cytometry. The production of platelet GPⅠbα shedding was detected by Western blot. The nonradioactive detection was used to measure PKC activity. Results PMA-induced platelet GPⅠbα shedding was completely inhibited by PKC inhibitor BIM or ROS antagonist dithiothreitol(DTT), respectively. PMA did dependently elevate ROS levels. PMA-induced ROS production was inhibited by PKC inhibitor and NAD(P)H oxidase inhibitor, but not mitochondrial ROS antagonist. BIM inhibited PMA-induced PKC activity, however, DTT did not. Conclusions PMA-induced platelet GPⅠbα shedding could be regulated by PMA-PKC-NAD(P)H oxidase-ROS-ADAM17-GPⅠbα signaling pathway.
Keyword:
Platelet; GPⅠbα shedding; Phorbol 12-myristate-13-acetate; Protein kinase C; Reactive oxygen species
引言
血小板在活化的同时,伴随有黏附受体GPⅠbα的酶切,产生的酶切片段称为糖盏蛋白(glycocalicin, GC)[ 1]。GPⅠbα酶切是血小板储存损伤(platelet storage lesion, PSL)一个重要的生物标志物[ 2]。因此,阐明GPⅠbα酶切的机制将有助于认识血小板储存损伤的机理。
佛波酯(phorbol 12-myristate-13-acetate,PMA)是常用的蛋白激酶C(protein kinase C, PKC)的活化剂。PMA能诱导GPⅠbα酶切[ 3],但是PMA诱导GPⅠbα酶切的分子机制尚未完全阐明。我们主要探讨了PMA诱导GPⅠbα酶切的分子机制。
材料和方法
一、材料
1. 研究对象 选取健康志愿者10名,男、女各5名,经上海中医药大学附属市中医医院伦理审查委员会批准同意,志愿者均签署知情同意书。
2. 主要试剂 抗GPⅠbα N端单克隆抗体SZ-2、辣根过氧化物酶(HRP)标记的羊抗小鼠IgG二抗购自Santa Cruz公司;GM6001购自Calbiochem公司;二硫苏糖醇(DTT)、NAD(P)H氧化酶抑制剂(DPI)、蛋白激酶C(PKC)抑制剂(BIM)、佛波酯(PMA)、ROS检测探针DCFDA购自Sigma公司;PKC活性检测试剂盒购自Promega公司;线粒体活化氧(ROS)拮抗剂MitoQ购自苏州沃盛化学有限公司。
二、方法
1. 制备洗涤血小板 取健康自愿者静脉血,用柠檬酸盐缓冲液(ACD)按照1∶7抗凝,380× g离心20 min得到富含血小板血浆(PRP),PRP经1 500× g离心20 min,沉淀用葡萄糖柠檬酸盐缓冲液(CGS)缓冲液悬浮,经离心、洗涤,最后用MTB液重悬,得到洗涤血小板,调整血小板浓度为3×108/mL,室温静置1 h使其恢复至静息状态以备用[ 4]。
2. Western blot检测GPⅠbα酶切片段 3×108个/mL洗涤血小板与1.0 μmol/L PMA或二甲基亚矾(DMSO) 37 ℃孵育30 min,2 600× g离心5 min,得到上清,加入5×上样缓冲液和β-巯基乙醇,最后样品经western blot检测GPⅠbα酶切片段。抑制实验:3×108个/mL 洗涤血小板预先与100 μmol/L BIM、DTT(25、 50、100、200 μmol/L)或DMSO 37 ℃孵育15 min后,再与1.0 μmol/L PMA 37 ℃孵育30 min。
3. 流式细胞仪检测ROS 3×108个/mL洗涤血小板分别与不同浓度PMA(0.25、0.5、1.0、2.0 μmol/L)或DMSO 37 ℃孵育30 min,加ROS检测探针DCFDA(20 μmol/L)孵育10 min,经稀释后,用流式细胞仪进行检测。抑制实验:3×108个/mL洗涤血小板预先与10 μmol/L DPI、100 μmol/L MitoQ、100 μmol/L BIM、200 μmol/L DTT或DMSO 37 ℃孵育15 min,再与1.0 μmol/L PMA 37 ℃孵育30 min。
4. 检测PKC活性 3×108个/mL洗涤血小板与1.0 μmol/L PMA或DMSO 37 ℃孵育30 min。抑制实验:3×108个/mL 洗涤血小板预先与200 μmol/L DTT、100 μmol/L BIM或DMSO 37 ℃孵育15 min,再与1.0 μmol/L PMA 37 ℃孵育30 min。使用非同位素标记的PKC活性检测试剂盒,先抽提PKC,再检测。具体步骤按说明书操作。
三、统计学方法
采用SPSS软件对实验数据进行统计分析,实验数据以 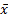
结果
一、BIM抑制PMA诱导的GPⅠbα酶切的分析
PMA是常用的PKC活化剂,能诱导GPⅠbα酶切。为了证实PKC在PMA诱导GPⅠbα酶切中的作用,本研究使用PKC特异性抑制剂BIM,同时使用解离素-金属蛋白酶17(a disintegrin and metalloproteinase 17, ADAM17)抑制剂GM6001作为对照。 图1结果显示,BIM完全抑制PMA诱导的GPⅠbα酶切;GM6001能完全抑制PMA诱导的GPⅠbα酶切。
 | 图1 BIM完全抑制PMA诱导的GPⅠbα酶切 |
二、DTT抑制PMA诱导的GPⅠbα酶切的分析
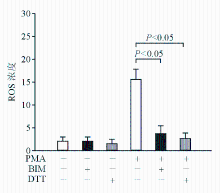
ROS通过氧化ADAM17的半胱氨酸残基调控ADAM17活性[ 5]。为了证实ROS是否参与PMA诱导的GPⅠbα酶切,本研究使用ROS抑制剂DTT。结果显示DTT浓度依赖地抑制PMA诱导的GPⅠbα酶切,在高浓度(200 μmol/L)时能完全抑制PMA诱导的GPⅠbα酶切。表明ROS参与调控PMA诱导的GPⅠbα酶切。见 图2。
 | 图2 DTT浓度依赖地抑制PMA诱导的GPⅠbα酶切 |
三、PMA浓度依赖地诱导血小板产生ROS
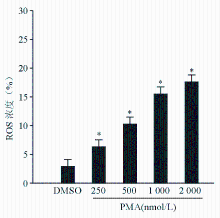
为了证实PMA是否诱导血小板产生ROS,本研究使用ROS检测探针DCFDA。 图3结果显示,PMA浓度依赖地诱导血小板产生ROS,表明PMA能够诱导血小板产生ROS。
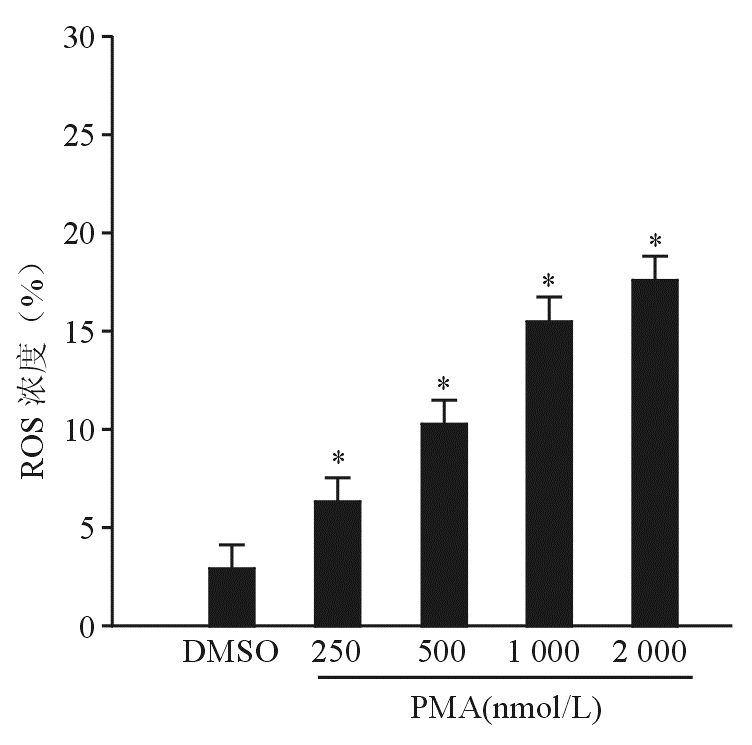 | 图3 PMA浓度依赖地诱导血小板产生ROS 注:与DMSO比较,* P<0.05 |
四、NAD(P)H氧化酶抑制剂抑制PMA诱导的ROS
ROS主要由线粒体呼吸链和NAD(P)H氧化酶产生[ 6]。为了证实PMA诱导产生的ROS的来源,本研究使用线粒体靶向的ROS拮抗剂MitoQ和DPI。 图4结果显示,MitoQ不抑制PMA诱导产生ROS,而DPI则抑制PMA诱导产生ROS,表明PMA诱导产生的ROS来自NAD(P)H氧化酶。
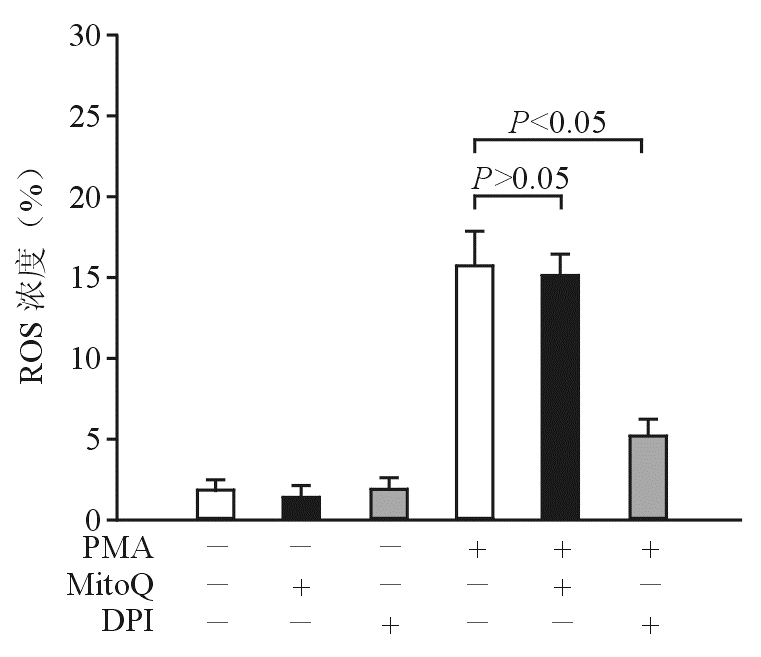 | 图4 DPI抑制PMA诱导的ROS |
五、PKC抑制剂抑制PMA诱导的ROS
为了证实PMA诱导产生ROS是通过活化PKC,本研究使用BIM。 图5结果显示,BIM抑制PMA诱导的ROS,表明PKC参与PMA诱导的ROS,提示PKC可能在ROS信号通路的上游。
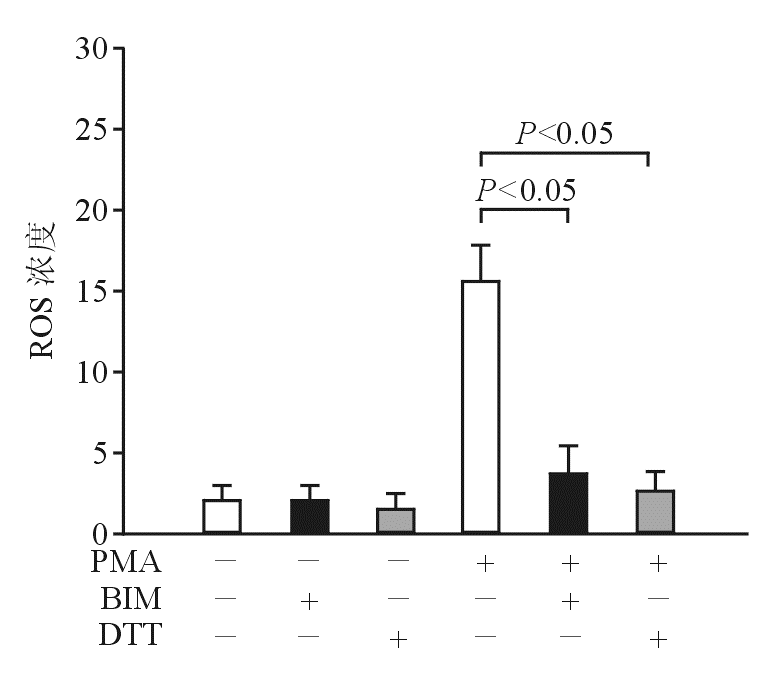 | 图5 BIM抑制PMA诱导的ROS |
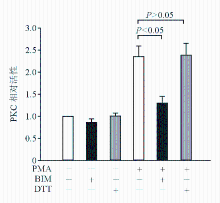
六、ROS抑制剂不抑制PMA诱导的PKC活化
为了进一步证实PKC与ROS之间的上下游关系,本研究使用PKC活性检测试剂盒。 图6结果显示,DTT不影响PMA诱导的PKC活性,而BIM则抑制PMA诱导的PKC活化。
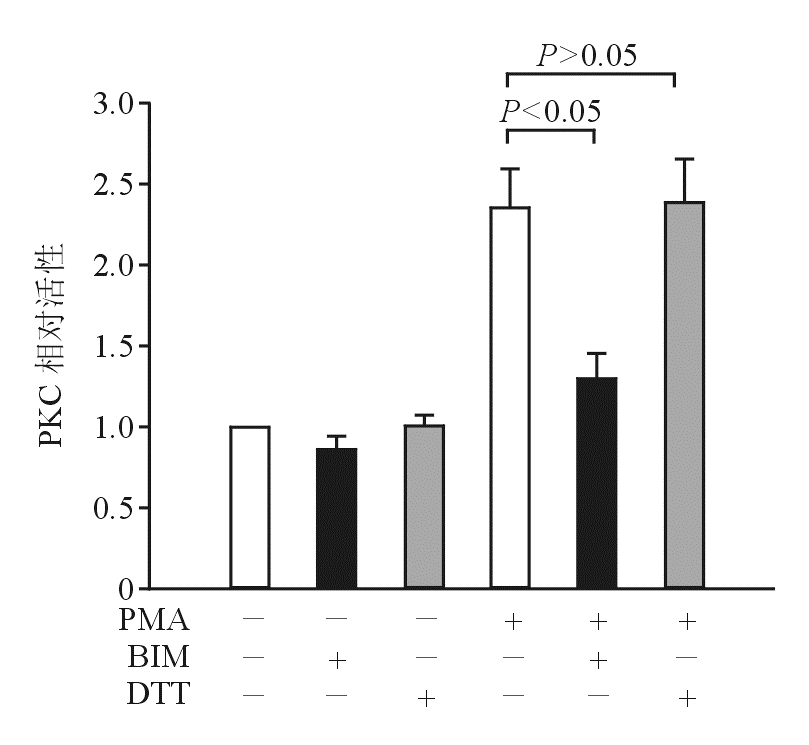 | 图6 DTT不抑制PMA诱导的PKC活化 |
讨论
GPⅠbα是血小板膜受体GPIb-IX-V复合物中最重要的一个亚基,能与血管性血友病因子(VWF)、凝血酶、P-选择素等结合,在血小板血栓形成的起始阶段起关键作用[ 7]。血小板在活化的同时,伴随有GPⅠbα酶切,产生的酶切片段称为GC[ 1]。
最近Bergmeier等[ 8]使用基因敲除小鼠证实ADAM17是负责酶切GPⅠbα的主要蛋白酶。ADAM17由824个氨基酸残基组成的I型跨膜蛋白,含有多个结构域。从N端开始包括:信号肽、前域、金属蛋白酶域、解离素域、表皮生长因子样域、半胱氨酸富含域、跨膜域和胞质尾[ 9]。已有报道,ADAM17的前域、半胱氨酸富含域、跨膜域和胞质尾参与ADAM17的活性调节[ 10]。其中,胞质尾在ADAM17活性调控中的调节存在争议,有些报道认为ADAM17胞质尾的丝氨酸被p38促分裂原活化蛋白激酶(p38MAPK)或PKC磷酸化引起ADAM17活性增加[ 11, 12];另一些报道认为胞质尾不参与调控ADAM17活性[ 13]。PMA是PKC活化剂,常用于ADAM17底物酶切的研究。Killock等[ 12]报道,PMA先活化PKC,活化的PKC直接磷酸化ADAM17胞质尾的丝氨酸,进而活化ADAM17引起底物的酶切。PMA能够诱导GPⅠbα酶切已被很多文献报道[ 3]。然而,PMA诱导GPⅠbα酶切的信号通路目前还未完全阐明。本研究使用各种抑制剂,结果表明在PMA诱导的GPⅠbα酶切中,可能存在“PMA-PKC-NAD(P)H氧化酶-ROS-ADAM17-GPⅠbα酶切”信号通路。本研究进一步支持ADAM17的胞质尾可能不参与调节ADAM17活性。NAD(P)H氧化酶是由5个主要亚基构成的酶复合体[ 14]。PKC如何引起NAD(P)H氧化酶活化本研究未涉及,可能通过直接或间接的方式,目前这方面的研究正在进行中。
近年来,ROS在调控ADAM17活性方面逐渐引起人们的关注[ 5]。ROS的来源主要有线粒体呼吸链和NAD(P)H氧化酶。本研究结果显示,PMA通过活化PKC,PKC通过直接或间接途径活化NAD(P)H氧化酶,产生ROS,ROS通过直接或间接的方式活化ADAM17,进而引起GPⅠbα酶切。有报道,ROS能直接通过氧化ADAM17的半胱氨酸残基直接活化ADAM17[ 5];也有报道,ROS通过活化p38MAPK进而引起ADAM17的活化[ 15]。ROS如何引起ADAM17活化本研究未涉及,可能通过直接或间接的方式,目前这方面的研究正在进行中。
最近的研究报道,钙离子依赖蛋白酶(calpain)参与A23187、凝血酶(在搅拌条件下)和calpain活化剂dibucaine诱导的GPⅠbα酶切[ 16]。本研究使用calpain抑制剂MDL28170,结果发现MDL28170不抑制PMA诱导的GPⅠbα酶切,表明不同的诱导剂引起的GPⅠbα酶切涉及不同的信号通路。
综上所述,本研究证实PMA诱导GPⅠbα酶切不是通过PKC直接活化ADAM17,而是通过NAD(P)H氧化酶产生的ROS活化ADAM17。本研究结果有助于进一步认识GPⅠbα酶切的机制。阐明GPⅠbα酶切的机制不仅有助于认识血小板血栓形成的机理,而且为认识血小板储存损伤提供新的视角。
The authors have declared that no competing interests exist.
参考文献
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|