{kind=link}
{kind=link}
{kind=link}
人结肠癌细胞中TGF-β1基因-509 C>T单核苷酸多态性对启动转录活性的影响
引用本文
顾星, 季鑫, 王爱华, 高春芳. 人结肠癌细胞中TGF-β1基因-509 C>T单核苷酸多态性对启动转录活性的影响[J].检验医学, 2015,25(9): 678-682
GU Xing, JI Xin, WANG Aihua, GAO Chunfang. Influence of -509 C>T single-nucleotide polymorphism on the transcriptional activity of transforming growth factor-beta 1 gene in human colorectal cancer cells[J]. Labratory Medicine, 2015,25(9): 678-682
Permissions
GU Xing, JI Xin, WANG Aihua, GAO Chunfang. Influence of -509 C>T single-nucleotide polymorphism on the transcriptional activity of transforming growth factor-beta 1 gene in human colorectal cancer cells[J]. Labratory Medicine, 2015,25(9): 678-682
Copyright©2010, 《检验医学》编辑部
《检验医学》编辑部
人结肠癌细胞中TGF-β1基因-509 C>T单核苷酸多态性对启动转录活性的影响
作者简介:顾 星,女,1981年生,博士,助理研究员,主要从事消化道疾病相关的基础研究。
通讯作者:高春芳,联系电话:021-81875131。
摘要
目的 探讨在人结肠癌细胞中转化生长因子β1 (TGF-β1)基因上游启动子-509 C>T位点多态性对其转录活性的影响。方法 以特定-509 C>T基因型患者DNA为模板,用聚合酶链反应(PCR)扩增得到1对长度为2.14 kb(-1 328~+812)含有-509 C>T 变异的TGF-β1上游基因片段,并将其与不含启动子的pCAT3-enhancer报告基因载体重组, 构建重组体phTGF 2.14C 和phTGF 2.14T。用脂质体转染法将2种重组体分别转染至人结肠癌细胞(SW480和LoVo细胞)中, 酶联免疫吸附试验(ELISA)测定转染细胞的报告基因CAT活性。结果 在SW480和LoVo细胞中,转染重组体phTGF 2.14C细胞的CAT活性均明显高于转染重组体phTGF 2.14T细胞的CAT活性( P<0.05)。结论 -509位点C等位基因可明显增强TGF-β1基因上游调控序列的转录活性。
关键词:
单核苷酸多态性; 转化生长因子β1; 启动子; 转录活性; 结肠癌
中图分类号:Q503
文献标志码:A
文章编号:1673-8640(2010)09-0678-05
Influence of -509 C>T single-nucleotide polymorphism on the transcriptional activity of transforming growth factor-beta 1 gene in human colorectal cancer cells
Abstract
Objective To study the influence of -509 C>T polymorphism on the transcriptional activity of transforming growth factor-beta 1 (TGF-β1) gene in human colorectal cancer cells.Methods Sequence -1 328-+812 (2.14 kb) of TGF-β1 gene containing the -509 C>T was selected as putative promoter by polymerase chain reaction (PCR) amplification.The -1 328-+812 of TGF-β1 gene and pCAT3-enhancer gene were recombined, and the phTGF 2.14C and phTGF 2.14T were constructed and transfected into human colorectal cancer cells (SW480 and LoVo) by liposomal transfection method. The transfected colorectal cancer cells were collected to analyze the reporter gene CAT activity by enzyme-linked immunosorbent assay (ELISA).Results In SW480 and LoVo cells, reporter gene CAT activity in cells transfected with phTGF 2.14C was significantly higher than that transfected with phTGF 2.14T ( P<0.05).Conclusions C allele at -509 C>T can increase the promoter activity of TGF-β1 gene in colorectal cancer cells.
Keyword:
Single-nucleotide polymorphism; Transforming growth factor-beta 1; Promoter; Transcriptional activity; Colorectal cancer
结肠癌是我国常见的消化道恶性肿瘤, 其发病率有逐年上升的趋势。转化生长因子β 1 (transforming growth factor-beta 1, TGF-β 1)是一种多功能多肽, 作为TGF-β 细胞因子超家族的成员调节多种靶基因的表达, 在肠上皮细胞的分化、移行、增殖及凋亡中发挥着重要作用[1]。血清中TGF-β 1升高与结肠癌进展、转移都显著相关。TGF-β 1基因序列上的SNP (single nucleotide polymorphism)与结肠癌相关性的研究也已有报道, 但结果并不一致。本研究在多年研究TGF-β 1转录活性及调控区SNP的基础上[2], 探寻TGF-β 1 基因上游-509 C> T多态性与结肠癌发生、发展的相关性, 从功能上探讨该位点多态性在结肠癌细胞中对调控转录作用的影响, 旨在明确结肠癌的发生、发展是否与该位点调控TGF-β 1转录活性有关。
材料和方法
一、材料和试剂
人结肠癌LoVo细胞和SW480细胞购自中科院细胞库, 由本实验室传代培养。培养基分别为10%胎牛血清(PAA, 德国)的F-12培养基和L-15培养基(Gibco, 美国), 0.25%胰蛋白酶消化传代(博光科技); DNA相对分子质量标记DL2000(TAKARA, 日本); 限制性内切酶Mlu Ⅰ 、Hind Ⅲ (TAKARA, 日本); T4 DNA连接酶(Promega, 美国); 质粒抽提纯化试剂盒plasmid midi kit (Qiagen, 德国); 小量割胶回收纯化试剂盒(华舜生物公司); DNA聚合酶KOD-plus(TOYOBO, 日本); 脂质体转染试剂lipofectamine 2000(Invitrogen, 美国); 载体质粒pCAT3-enhancer(Promega, 美国)、β -半乳糖苷酶表达质粒pSVβ -gal(Promega, 美国)和含氯霉素乙酰基转移酶(CAT)的表达质粒pCAT6.2(Promega, 美国); 细胞裂解缓冲液(Roche, 德国); BCA蛋白定量试剂盒(Pierce, 美国); 酶标仪(Thermo, 美国); 聚合酶链反应(RCR)仪Rotter Gene RG-3000(Corbett Research, 澳大利亚)。
二、方法
1. 构建重组质粒 载体质粒pCAT3-enhancer不含启动子, 但含有SV40增强子的CAT报告基因。重组体phTGF 2.14以特定-509 C> T基因型患者DNA为模板, 采用PCR扩增得到1对长度为2.14 kb含有-509 C> T位点的TGF-β 1上游基因片段, 相应于TGF-β 1基因上游-1 328~+812序列, 分别构建phTGF 2.14C、phTGF 2.14T, 其上游引物序列为5'-GATTCGACGCGTAGATCACTTTG-GCTGCTGT-3', 下游引物序列为5'-TAGACC-AAGCTTGAGCGCGAACAGGGC-3'。两端引物分别含6个保护碱基和限制性内切酶Mlu Ⅰ 、Hind Ⅲ 识别位点(下划线部分)。引物由上海基康生物技术公司合成。扩增产物经割胶小量柱纯化回收后, 用限制性内切酶Mlu Ⅰ 、Hind Ⅲ 酶切并与相同黏末端的线性pCAT3-enhancer Vector经T4 DNA连接酶连接, 连接混合物转化至感受态大肠埃希菌JM109并筛选, 小量酶切, 经测序(上海基康生物技术公司)鉴定正确后, 大量扩增阳性克隆、抽提、纯化质粒重组体, 紫外分光光度计测得率、纯度。
2. DNA 转染 转染采用脂质体转染试剂, 操作按照试剂说明进行。具体方法如下:结肠癌LoVo和SW480细胞以105/孔接种于12孔培养板, 待细胞80%融合生长, 换无血清培养基。按DNA与脂质体转染试剂为1∶ 2.5的比例将重组质粒1 μ g与转染试剂复合物加入上述细胞培养孔中, 继续培养。同时另转染β -半乳糖苷酶表达质粒(pSVβ -gal)1 μ g作为内对照及含CAT的表达质粒(pCAT6.2)作阳性对照。6 h后换含10%胎牛血清培养基继续培养48 h。上述实验采用4复孔, 并重复3次。
3. CAT 活性测定及蛋白定量、β -gal活性 终止培养的细胞用预冷的磷酸盐缓冲液(PBS)洗3次, 以裂解缓冲液裂解细胞后, 取细胞裂解液进行蛋白定量、CAT-酶联免疫吸附试验(ELISA)定量。β -gal活性测定方法如下:100 μ L细胞裂解液加入100 μ L β -gal assay 2× buffer混匀, 置37 ℃12 h后加入1 mol/L碳酸钠混匀, 酶标仪405 nm/490nm 双波长测定吸光度(A)值。
三、统计学方法
应用SPSS 11.0软件对研究数据进行统计分析。各指标用
结 果
一、重组体鉴定
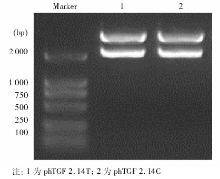
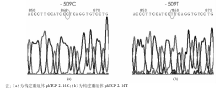
以特定-509 C> T基因型患者DNA为模板, 用PCR扩增得到1对长度为2.14 kb(-1 328~+812)含有-509 C> T变异的TGF-β 1上游基因片段, 并将其与不含启动子的pCAT3-enhancer 载体重组, 构建重组体phTGF 2.14C和phTGF2.14T。经酶切后电泳初步鉴定正确, 见图1, 即载体为4.3 kb的pCAT3-enhancer, 插入的启动子片段长度为2.14 kb, 进一步经测序验证插入片段分别含有-509 C> T变异的TGF-β 1上游基因片段, 并与GenBank相应序列一致, 见图2。
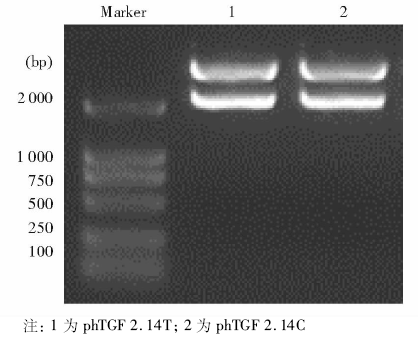 | 图1 2种重组质粒酶切后电泳结果 |
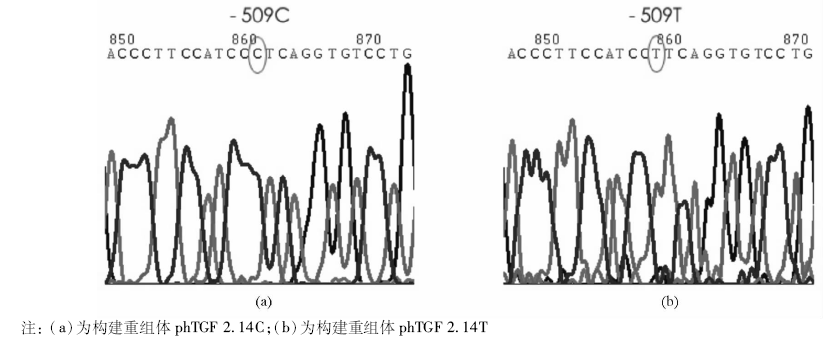 | 图2 构建重组体phTGF 2.14C和phTGF 2.14T测序验证结果 |
二、转染重组体的细胞报告基因表达水平
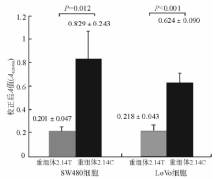
结肠癌细胞在转染不同重组体48 h后, ELISA测定细胞CAT表达量, 同时测定细胞裂解液蛋白浓度。CAT测定结果用相应蛋白含量及β -gal活性校正以保证在相同的细胞量及转染效率前提下比较报告基因活性。结果见图3。
1. 在SW480细胞中, 转染重组体phTGF 2.14C的细胞测得平均蛋白浓度为380 μ g/mL, 转染重组体phTGF 2.14T细胞测得平均蛋白浓度为420 μ g/mL。根据蛋白浓度校正后, 得到转染重组体phTGF 2.14C的CAT活性对应的平均A值为0.829, 转染重组体phTGF 2.14T细胞的平均A值0.201, s值分别为0.243和0.047, 转染重组体phTGF 2.14C细胞的CAT活性明显高于转染重组体phTGF 2.14T细胞的CAT活性(t =5.063, P=0.012)。重复检测3次, 结果类似。见图3。
2. 在LoVo细胞中, 转染重组体phTGF 2.14C的细胞测得平均蛋白浓度为223 μ g/mL, 转染重组体phTGF 2.14T细胞测得平均蛋白浓度为244 μ g/mL。根据蛋白浓度校正后, 得到转染重组体phTGF2.14C细胞的平均A值为0.624, 转染重组体phTGF 2.14T细胞的平均A值为0.218, s值分别为0.090和0.043, 转染重组体phTGF 2.14C细胞的CAT活性明显高于转染重组体phTGF 2.14T细胞的CAT活性 (t=8.155, P< 0.001)。重复检测3次, 结果类似。见图3。
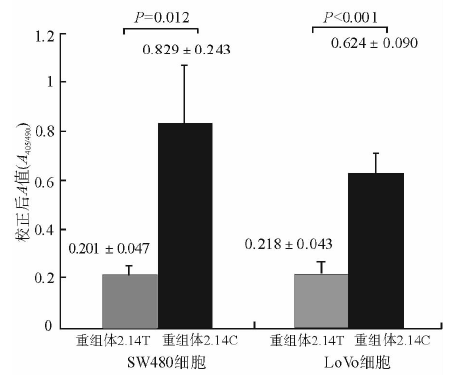 | 图3 转染重组体的结肠癌细胞CAT活性测定 |
讨 论
TGF-β 1具有多种广泛功能, 在多种疾病的发生、发展中地位特殊。国外及本实验室的研究发现, TGF-β 1在肿瘤、脏器纤维化、创伤修复、炎症性疾病及自身免疫性疾病等发生、发展中具有重要意义[3, 4]。
在TGF-β 1常见的8个SNP位点中, 中国人在-800、-988位点及codon 25、codon 263基本不存在基因多态性, 常见的变异主要存在于-509 C> T和codon 10(Leu> Pro), 其中-509存在于TGF-β 1转录起始位点上游启动子区域, 多年来已发现该位点与多种疾病的发生、发展有关[5]。
前期本课题组围绕TGF-β 1基因启动子区域-509 C> T多态性与人类多种疾病的相关性进行了详尽的研究。我们已发现该位点与慢性乙型肝炎患者的肝细胞癌易感性显著相关, C等位基因携带者具有更高的发病风险[6]。在与结肠癌相关性方面, 我们通过病例-对照实验, 对150例结肠癌病例和500名正常对照者该位点多态性进行了分析, 结果发现其与被测人群结肠癌的发生以及进程不具有显著相关性[7], 这与Wu等[8]2009年在高加索人群中得到的检测结果一致。而今年另一篇报道表明, 在伊朗人群中-509 C> T多态性与结肠癌易感性显著相关, T等位基因具有保护作用[9]。为进一步弄清这一位点变异对于结肠癌易感性可能带来的影响, 我们采用计算机软件模拟分析其核转录因子结合情况, 分析了以-509为核心的上、下游60 bp的序列即-449~-570 bp的DNA结合蛋白, 发现-509 C> T的变化, 相应核蛋白转录因子的结合谱是不同的[10]。这一结果提示-509 C> T的变化理论上可影响核转录因子的结合模式, 从而影响其转录水平。
SW480和LoVo细胞是美国标准菌库收录的2种常用的人结肠癌细胞株, 多年来被广泛用于人结肠癌发生、发展机制相关的细胞学研究。在本实验中, 我们通过构建含报告基因且启动子区域为含该多态性位点序列的重组体, 分别转染至这2种结肠癌细胞中, 由报告基因所测活性反映出-509 C> T造成了转录调控水平的差异。我们的结果表明, -509位点C等位基因表达TGF-β 1的活性明显高于T等位基因, 提示C等位基因可明显增强TGF-β 1基因上游调控序列的转录活性。这与本实验室前期在肝硬化中对于TGF-β 1基因该位点的研究结果[11]一致。
已有研究表明血清TGF-β 1升高与结肠癌进展、转移显著相关, TGF-β 1升高患者较对照组复发风险升高。也有报道表明TGF-β 1可作为反映结肠肿瘤侵袭性和预后的参考指标之一。在结肠癌发生早期, TGF-β 1作为肿瘤抑制因子起作用, 通过G期到S期抑制正常细胞生长和肿瘤细胞发生。此外, TGF-β 1也可以刺激其他生长因子如血小板衍化生长因子(PDGF)、碱性成纤维细胞生长因子(bFGF)和血管内皮生长因子(VEGF)的分泌, 这些生长因子能促进TGF-β 1诱导的细胞增殖和血管形成。所以, 我们推测-509 C> T多态性位点T等位基因变异在结肠癌细胞中抑制了TGF-β 1的转录活性, 降低了TGF-β 1的表达量, 从而对肿瘤的发展和进程具有一定的保护作用。
本研究将有助于我们进一步深入认识TGF-β 1基因中不同位点多态性对TGF-β 1转录调控、分泌和功能的影响。研究这些基因多态性与疾病的相关性, 阐述TGF-β 1基因结构对其蛋白表达及功能的影响, 必将为结肠癌的诊断、治疗与逆转提供新的思路。
The authors have declared that no competing interests exist.
参考文献
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|