{kind=link}
{kind=link}
儿童肠球菌多重耐药与I类整合子的检测
引用本文
吕萍, 徐樨巍, 宋文琪, 董方, 杨永弘, 沈叙庄. 儿童肠球菌多重耐药与I类整合子的检测. 2009, 24(3): 219-222
LÜ Ping, XU Xiwei, SONG Wenqi, DONG Fang, YANG Yonghong, SHEN Xuzhuang. Multidrug resistance of Enterococcal isolates in pediatric and test of class Ⅰ integron . Labratory Medicine, 2009, 24(3): 219-222
Permissions
LÜ Ping, XU Xiwei, SONG Wenqi, DONG Fang, YANG Yonghong, SHEN Xuzhuang. Multidrug resistance of Enterococcal isolates in pediatric and test of class Ⅰ integron . Labratory Medicine, 2009, 24(3): 219-222
Copyright©2009, 《检验医学》编辑部
《检验医学》编辑部
儿童肠球菌多重耐药与I类整合子的检测
作者简介:吕 萍,女,1971年生,硕士,主管技师,主要从事病原微生物研究。
摘要
目的
了解儿童临床分离肠球菌的耐药特征及其多重耐药与I类整合子的相互关系。
方法采用琼脂稀释法测定常用抗菌药物对152株肠球菌的最低抑菌浓度(MIC),用聚合酶链反应(PCR)检测肠球菌I类整合子和I类整合酶基因。
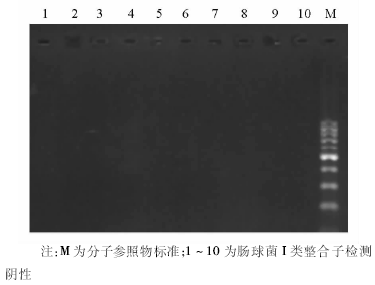
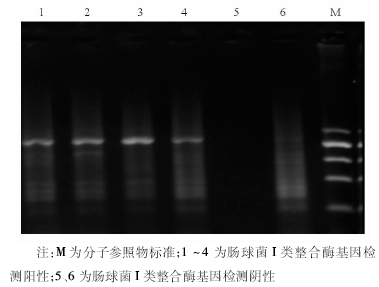
结果屎肠球菌对氨苄西林、阿莫西林-克拉维酸、环丙沙星的耐药率分别为96.8%、95.2%和84.1%,粪肠球菌对上述3种抗菌药物的耐药率分别为23.6%、18.0%和49.4%,屎肠球菌的耐药率明显高于粪肠球菌( P<0.001);粪肠球菌中有2株对万古霉素的MIC为8 μg/mL,粪肠球菌和屎肠球菌对替考拉宁均敏感。儿童多重耐药肠球菌发生率高达93.7%。屎肠球菌耐药模式以耐氨苄西林、红霉素、环丙沙星、利福平、四环素、高水平庆大霉素6种抗菌药物为主,占屎肠球菌的59%,粪肠球菌以耐四环素、红霉素、利福平、氯霉素、高水平庆大霉素5种抗菌药物为主,占粪肠球菌的26%。全部152株肠球菌未检测到I类整合子,仅有5株检测到I类整合酶基因。
结论儿童肠球菌多重耐药十分严重,儿童肠球菌多重耐药与I类整合子和I类整合酶基因尚无明显关系。
关键词:
肠球菌; 整合酶基因; 整合子; 多重耐药
中图分类号:R378.1
文献标志码:A
文章编号:1673-8640(2009)03-0219-04
Multidrug resistance of Enterococcal isolates in pediatric and test of class Ⅰ integron
Abstract
Objective
To explore the resistant characteristic and the correlation between multidrug resistance and class I integron in Enterococcal isolates from children.
MethodsMininal inhibitory concentrations (MIC) of 152 Enterococcal isolates were determined by agar dilution. Class I integron and class I integrase were detected by polymerase chain reaction (PCR).
ResultsThe resistant rate of Enterococci faecium ( E.faecium) to ampicillin,amoxicillin-clavulanic acid,ciprofloxacin was 96.8%,95.2% and 84.1% respectively. The resistant rate of E.faecalis to ampicillin,amoxicillin-clavulanic acid,ciprofloxacin was 23.6%,18.0% and 49.4% respectively. The resistant rates of E.faecium to ampicillin,amoxicillin-clavulanic acid,ciprofloxacin were higher than E.faecalis( P<0.001). The MIC of vancomycin was 8 μg/mL in two E.faecalis. E.faecalis and E.faecium isolates were all susceptible to teicoplanin. The multiresistant rate to antibiotic in the Enterococci was 93.7%. The patterns of multiresistance to antibiotic in E.faecium were ampicillin,erythromycin,tetracycline,ciprofloxcin,rifampin,high level gentamicin.The patterns of multiresistance to antibiotic in E.faecalis were tetracycline,erythromycin,rifampin,chloramphenicol,high level gentamicin. Class I integron was negative among all the Enterococcal isolates,only five Enterococcal isolates were tested class I integrase.
ConclusionsThe antimicrobial resistant status of the Enterococci is very serious.The multidrug resistance has no correlation to class I integron and class I integrase.
Keyword:
Enterococci; Integrase; Integron; Multidrug resistance
目前, 由具有捕获及表达外来耐药基因盒能力的整合子系统介导的细菌耐药机制越来越引起人们的关注[1], 其对多重耐药性研究有着非常重要的意义[2]。有研究表明, 细菌整合子可以携带6种或更多耐药基因盒[3, 4], 并同时对十几种抗菌药物产生耐药性, 某些质粒携带的耐药基因也与整合子有关[5, 6], 由于细菌可以不断地通过整合酶基因从周围环境捕获耐药基因盒, 而使其具有多重耐药性。同时, 整合子可以通过存在于质粒、转座子等的可移动基因元件, 在同种或不同种属之间进行基因的水平转移, 从而使细菌的耐药性得以传播。众多文献报道整合子介导的耐药在革兰阴性细菌中很普遍, 在革兰阳性细菌中存在的情况报道甚少, 因此, 本研究旨在探讨儿童肠球菌的多重耐药与I类整合子的相互关系及I类整合子在肠球菌耐药中的作用。
材料和方法
一、材料
1. 菌株来源 受试菌为2004年7月至2005年11月北京儿童医院门诊和住院患儿临床标本中分离的病原菌共152株, 同一患者重复菌株选择首次分离菌。质控菌由卫生部临床检验中心提供, 包括粪肠球菌(ATCC 29212)、粪肠球菌(ATCC 51299)。
2. 抗菌药物 氨苄西林、阿莫西林-克拉维酸、红霉素、四环素、利福平、环丙沙星、氯霉素、万古霉素、庆大霉素、链霉素均为中国药品生物制品检定所提供; 替考拉宁购自国药集团药业有限公司。
3. 培养基及其他试剂 水解酪蛋白胨(MH)琼脂和脑心浸液(BHI)琼脂、头孢硝噻吩Nitrocefin纸片为英国Oxoid公司产品。聚合酶链反应(PCR)试剂和Ladder Marker均由赛百胜基因技术有限公司提供。
二、方法
1. 药敏试验 最低抑菌浓度(MIC)的测定采用琼脂稀释法, 严格按照美国临床实验室标准化研究所(CLSI)推荐的琼脂稀释法进行操作和判定结果[7]。庆大霉素、链霉素分别以MIC> 500 μ g/mL、MIC> 1 000 μ g/mL为高度耐药的界值。35 ℃孵箱孵育18~20 h, 万古霉素、高浓度庆大霉素和链霉素为24 h。高浓度庆大霉素和链霉素每次实验均含标准菌株粪肠球菌(ATCC 29212)和粪肠球菌(ATCC 51299)做质控监测。
2. 细菌DNA模板的提取 采用煮沸法[8]。
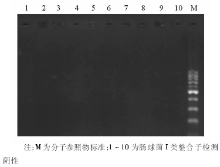
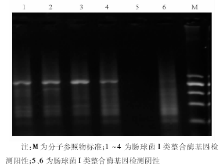
3. PCR I类整合子和I类整合酶基因特异引物序列为5'-CS:5'-GGCATCCAAGCAAG-3'; 3'-CS:5'-AAGCAGACTTGACCTGA-3'; IntI1L:5'-ACATGTGATGGCGACGCACGA-3'; IntI1R:5'-ATTTCTGTCCTGGCTGGCGA-3'。 PCR扩增:50 μ L PCR反应体系。TaqDNA聚合酶2 U, 10× 缓冲液(500 mmol/L KCl, 100 mmol/L Tris-HCl, 1.0%Triton X-100, 250 mmol/L MgCl2)5 μ L, dNTPs各200 mmol/L, 上样染料和稳定剂各5 μ L, 模板5 μ L, 引物为20 pmol。扩增条件:94 ℃预变性5 min, 94 ℃变性30 s, 57 ℃退火30 s, 72 ℃延伸1 min, 30个循环, 再72 ℃延伸10 min。PCR产物分析:1%琼脂糖凝胶电泳后, 在紫外光源下观察结果, 用Ladder Marker作参照。
三、统计学方法
数据用WHONET5.3软件进行分析处理, SPSS 11.5软件进行耐药率之间χ 2检验, 以P< 0.05为差异有统计学意义。
结 果
一、肠球菌常用抗菌药物MIC的测定
89株粪肠球菌对替考拉宁全部敏感, 有2株对万古霉素的MIC50为8 μ g/mL, 为万古霉素中介株。对红霉素、四环素和利福平的耐药率分别为93.3%、79.8%和71.9%。63株屎肠球菌对替考拉宁、万古霉素全部敏感, 而对其他抗菌药物均为高度耐药。屎肠球菌对氨苄西林、阿莫西林-克拉维酸、环丙沙星的耐药率分别为96.8%、95.2%和84.1%, 明显高于粪肠球菌(P均< 0.001); 对利福平、高浓度庆大霉素和链霉素的耐药率也高于粪肠球菌(P均< 0.05), 而对氯霉素的耐药率明显低于粪肠球菌(P=0.000)。见表1。
| 表1 89株粪肠球菌和63株屎肠球菌的MIC及耐药率 |
二、肠球菌的多重耐药分析
在152株肠球菌中有140株对3种和3种以上抗菌药物耐药, 占92.0%; 对4种以上抗菌药物耐药的肠球菌有125株, 占82.0%; 对5种以上抗菌药物耐药的肠球菌有102株, 占67.0%; 对6种以上抗菌药物耐药的肠球菌有73株, 占48.0%。屎肠球菌以耐氨苄西林、红霉素、环丙沙星、利福平、四环素、高水平庆大霉素6种抗菌药物为主, 占屎肠球菌的59%, 在屎肠球菌中联合氨苄西林、红霉素、环丙沙星、利福平4种抗菌药物耐药的约占屎肠球的90%; 而粪肠球菌则以红霉素、利福平、四环素、氯霉素、高水平庆大霉素为主, 占粪肠球菌的26%, 见表2。由此可见, 儿童肠球菌的多重耐药十分严重, 且多重耐药株明显高于成年人[9]。
| 表2 肠球菌对抗菌药物常见联合耐药摸式 |
讨 论
肠球菌是人类和动物消化道寄生的革兰阳性共栖菌。近年来, 由于广谱抗菌药物和免疫抑制剂的广泛使用, 使本来就对某些抗菌药物固有耐药的肠球菌耐药性越来越严重。目前, 已证实对一种或多种抗菌药物耐药的肠球菌临床分离率在不断增加[10]。89株粪肠球菌对替考拉宁全部敏感, 出现2株万古霉素中介株, 63株屎肠球菌对替考拉宁、万古霉素全部敏感, 而对其他抗菌药物均为高度耐药, 屎肠球菌对氨苄西林、阿莫西林-克拉维酸、环丙沙星、利福平、高浓度庆大霉素和链霉素的耐药率高于粪肠球菌, 而对氯霉素的耐药率则明显低于粪肠球菌。肠球菌的多重耐药非常严重, 尤以屎肠球菌更为严重, 因此临床应根据粪肠球菌和屎肠球菌的不同耐药性选择抗菌药物。
肠球菌除了对多种抗菌药物天然耐药以外, 主要是通过染色体以外的遗传单位如质粒、转座子等携带耐药基因从而对氨基糖苷类、糖肽类、β -内酰胺类、四环素类、大环内酯类抗菌药物耐药。整合子-基因盒系统于1989年由Hall等[11]正式提出, 是细菌的天然克隆与表达系统, 能捕获外来耐药基因, 在整合子中可形成多种耐药基因的组合、排列, 整合于整合子上的基因盒可借助整合子的强启动子而得以表达, 是近年来发现的细菌耐药性传播的机制之一。临床菌株中发现的多为I类整合子, 携带β -内酰胺类、氨基糖苷类、甲氧磺胺嘧啶类耐药基因, 常存在于在肠杆菌[12]、假单胞菌和鲍曼不动杆菌中。近年来国内有报道肠球菌中发现I类整合子, 且粪肠球菌和屎肠球菌I类整合子携带率分别为53.1%和51.5%[13], 而本实验全部152株肠球菌未检测到I类整合子基因, 说明I类整合子在我院分离的肠球菌中并不常见, 这与国内李爽等[14]报道一致。由此可以看出, I类整合子基因与肠球菌的多重耐药在不同医院分离菌株中存在差异。尽管我院肠球菌多重耐药十分严重, 但肠球菌的多重耐药与I类整合子基因并无明显关系, 可能存在其他的作用机制。
The authors have declared that no competing interests exist.
参考文献
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|