{kind=link}
{kind=link}
{kind=link}
阿萨希毛孢子菌致急性心肌梗死患者肺部感染一例
引用本文
李继红, 赵建宏. 阿萨希毛孢子菌致急性心肌梗死患者肺部感染一例. 2009, 24(2): 159-160

Permissions
Copyright©2009, 《检验医学》编辑部
《检验医学》编辑部
阿萨希毛孢子菌致急性心肌梗死患者肺部感染一例
作者简介:李继红,女,1971年生,硕士,主管技师,主要从事病原微生物耐药及相关耐药基因研究。
关键词:
阿萨希毛孢子菌; 急性心肌梗死; 肺部感染
中图分类号:R379.2
文献标志码:B
文章编号:1673-8640(2009)02-0159-02
阿萨希毛孢子菌(Trichosporon asahii)原属皮肤毛孢子菌, 为酵母样真菌, 目前认为该菌为条件致病菌, 国外报道多见于血液病、恶性肿瘤、免疫缺陷、白细胞减少症等患者的皮肤感染[1, 2] 。我们于2007年3月发现了1例心肌梗死继发阿萨希毛孢子菌所致肺部感染的患者。
一、病例资料
患者男, 74岁, 既往有高血压史20年, 脑梗死3年, 4 d前间断性胸痛, 1 d前症状加重, 持续性胸痛伴大汗, 当地心电图诊断为急性心肌梗死, 抗凝治疗后, 为进一步诊治于2007年3月15日入我院急诊重症监护病房。诊断为:(1) 冠心病急性心肌梗死; (2) 高血压病; (3) 陈旧性脑梗死, 给予对症治疗。次日, 患者出现呼吸稍急促, 无胸痛胸闷, 两肺呼吸音粗, 可闻及较多湿性啰音, 间断咳嗽, 痰无力咳出。胸片显示, 右肺下野透光度减低, 考虑胸膜病变, 右侧膈肌位置抬高。白细胞10.8× 109/L, 中性粒细胞82.3%, 淋巴细胞5.2% , 单核细胞11.2 %, 嗜酸性粒细胞0.7%, 嗜碱性粒细胞0.6%, 血红蛋白 148 g/L, 血小板152× 109/L, 考虑存在肺部感染, 用氨苄西林常规剂量治疗。3月17日因症状无改善, 患者出现胸闷、气短、烦躁。给予插管呼吸机辅助治疗后病情相对平稳。送痰细菌涂片、痰培养。细菌涂片显示大量菌丝及孢子。痰细菌培养:阿萨希毛孢子菌纯生长。因该菌较少见, 于次日上午、下午又连续2次送检痰培养, 均为阿萨希毛孢子菌纯培养。为慎重起见, 我们于第3天对呼吸机的出口拭子、气管插管入口拭子以及通过患者气管插管抽取的痰标本进行了对照培养。结果为呼吸机的出口拭子、气管插管入口拭子均无菌生长, 而在通过患者气管插管抽取的痰标本中仍然有纯培养。血标本、尿标本培养均为阴性。根据药敏结果选用敏感的两性霉素B脂质体联合氟康唑进行治疗, 症状缓解。于治疗的第7天培养转为阴性。
二、结果
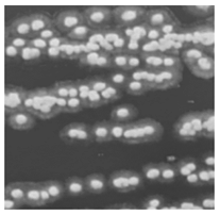
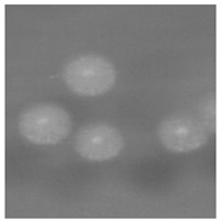
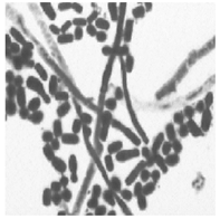
1. 培养 将待检痰标本分别接种于血琼脂平板、麦康凯平板、沙保弱培养基。37 ℃培养24 h, 血琼脂平板生长出白色、干燥、边缘不整齐、触之易碎的菌落, 见图1。麦康凯平板无生长。沙保弱培养基生长为圆形、扁平、白色中央浓集、边缘菲薄、毛玻璃样边缘、有菌丝生长的真菌相菌落, 见图2。转种科玛嘉显色培养基(CA), 37 ℃培养24 h后生长产生绿色色素的菌落。革兰染色后镜检可见大量菌丝、节孢子和多种形状的小分生孢子, 菌丝分枝分隔粗细不等, 节孢子数目不等、长短不一, 每一节孢子多呈矩形和椭圆形, 见图3。
 | 图1 血琼脂平板生长1d |
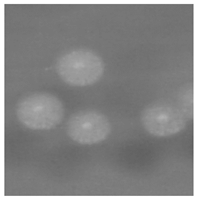 | 图2 沙保弱培养基生长1d |
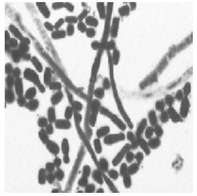 | 图3 显微镜下形态(× 1000) |
2. 生化鉴定 利用API 20C AUX 试剂盒检测, 结果提示该菌为阿萨希毛孢子菌, 鉴定百分率为99.9%, T=0.70。
3. 药敏结果 采用法国生物梅里埃公司真菌药敏定量鉴定卡FUNGS-2进行药物敏感实验, 结果为氟胞嘧啶敏感, 两性霉素B敏感, 伊曲康唑中介, 氟康唑耐药。
三、讨论
阿萨希毛孢子菌广泛存在于自然界中, 是一种机会致病菌, Sugita等[3]通过DNA序列分析认为皮肤表面的毛孢子菌是毛孢子菌病的主要致病菌, 该菌可引起夏季超敏性肺炎, 免疫力低下患者的播散性感染, 皮肤的多发紫癜样丘疹, 头部毛发的白结节病等。该患者既往有冬季咳嗽、咳痰病史, 但否认过敏性哮喘病史, 同时患者的症状、体征、各项检查指标均支持肺部感染诊断。应用两性霉素B脂质体7 d后再次培养阴性, 说明该菌为引起患者肺部感染的病原菌。患者脑梗死3年, 免疫力低下, 长期卧床, 室内空气的潮湿及不流通可能是造成患者感染的原因。
有报道, 该菌在37及25 ℃生长为不同菌落, 37 ℃生长快, 为奶油色的酵母样菌落, 25 ℃生长成乳白色至淡黄色, 我们把该菌在沙保弱培养基上不同温度下培养5 d后, 未发现该菌在同一培养基上的双相生长现象, 却观察到在血琼脂上生长始终为酵母型, 沙保弱培养基生长始终为真菌相, 对阿萨希毛孢子菌的此种生长现象, 有待于进一步探讨。
近年来, 随着免疫缺陷患者的增多, 应关注阿萨希毛孢子菌可能的感染, 杨蓉娅等[4]进行动物实验后发现两性霉素B脂质体联合氟康唑可有效抑制该菌在小鼠体内的感染和扩散, 但该菌对常用的咪唑类抗真菌药物常表现为无效, 甚至两性霉素B联合咪唑类抗真菌药物对于粒细胞缺乏患者的阿萨希毛孢子菌感染治疗失败率也高达100%[5], 因此, 对于该方案的应用有待于积累更多的研究资料。
基础免疫状态对阿萨希毛孢子菌感染患者的康复有较大的影响, 对于机体免疫力正常的患者, 即使体外耐药, 体内用药也可能是敏感的, 而对于免疫缺陷患者, 感染的治疗则需要严格依照药敏试验的结果进行。
The authors have declared that no competing interests exist.
参考文献
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|