{kind=link}
{kind=link}
{kind=link}
SCD1过表达影响高脂诱导鼠BRL肝细胞膜电位变化研究
引用本文
蔡德丰, 范建高, 陆元善, 蔡晓波. SCD1过表达影响高脂诱导鼠BRL肝细胞膜电位变化研究[J].检验医学, 2015,25(9): 688-692
CAI Defeng, FAN Jiangao, LU Yuanshan, CAI Xiaobo. Research on the overexpression of SCD1 affecting the change of mitochondrium membrane potential in rat BRL hepatocytes caused by fat-rich diet induction[J]. Labratory Medicine, 2015,25(9): 688-692
Permissions
CAI Defeng, FAN Jiangao, LU Yuanshan, CAI Xiaobo. Research on the overexpression of SCD1 affecting the change of mitochondrium membrane potential in rat BRL hepatocytes caused by fat-rich diet induction[J]. Labratory Medicine, 2015,25(9): 688-692
Copyright©2010, 《检验医学》编辑部
《检验医学》编辑部
SCD1过表达影响高脂诱导鼠BRL肝细胞膜电位变化研究
作者简介:蔡德丰,男,1979年生,学士,技师,主要从事脂代谢相关疾病研究。
通讯作者:陆元善,联系电话:021-63240090-4306。
摘要
目的 探讨硬脂酰辅酶A去饱和酶1(SCD1)过表达影响大鼠BRL肝细胞系凋亡作用的可能机制。方法 通过台盼兰染色检测不同浓度棕榈酸(PA)诱导BRL肝细胞死亡率确定下游实验PA添加浓度。培养细胞分组为非加药组和加药组。非加药组包括一般培养组(CON)、一般培养加阴性病毒组(NC)及一般培养加过表达病毒组(SCD1-LV);加药组包括一般培养加PA组(CON+)、一般培养加阴性病毒和PA组(NC+)及一般培养加过表达病毒和PA组(SCD1-LV+) 。实时荧光定量聚合酶链反应(PCR)检测SCD1 mRNA表达的变化,流式细胞术测细胞线粒体膜电位(MMP)的变化,用膜电位正常细胞百分比(JC-1+%)表示MMP改变。结果 与对照组相比,400 μmol/L PA诱导72 h细胞死亡率明显上升( P<0.01)。PA诱导引起CON+及NC+组SCD1表达下降,SCD1-LV+ 组及SCD1-LV组SCD1表达明显上升; CON+和NC+组JC-1+%较低,SCD1-LV+ 组JC-1+%与对照组相似。结论 用SCD1慢病毒载体感染BRL肝细胞系,SCD1表达升高,增强细胞对饱和脂肪酸的去饱和作用,过表达SCD1能降低PA诱导的脂毒性,使紊乱的MMP得到恢复。
关键词:
硬脂酰辅酶A去饱和酶1; 棕榈酸; 慢病毒载体; 线粒体膜电位
中图分类号:Q556
文献标志码:A
文章编号:1673-8640(2010)09-0688-05
Research on the overexpression of SCD1 affecting the change of mitochondrium membrane potential in rat BRL hepatocytes caused by fat-rich diet induction
Abstract
Objective To investigate the mechanism of stearoyl-CoA desaturase-1 (SCD1) overexpression affecting the apoptosis of rat BRL hepatocytes induced by a fat-rich diet.Methods The death rates of BRL hepatocytes under different palmitic acid (PA) concentrations were detected by Trypan blue staining. The concentration of PA was certained for the following determination. Cultured cells were classified into two groups (no-PA added group and PA added group). No-PA added group included cultured ordinary group(CON), CON plus negative virus cultured group(NC) and CON plus SCD1 overexpression virus cultured group(SCD1-LV). PA added group included PA plus cultured ordinary group(CON+), CON plus negative virus and PA cultured group(NC+), and CON plus SCD1 overexpression virus and PA cultured group(SCD1-LV+). SCD1 mRNA expression was detected by real time polymerase chain reaction (PCR). The changes of mitochondrium membrane potential (MMP) were detected by flow cytometry. The percentage of BRL hepatocytes of normal membrane potential (5, 5', 6, 6'-tetrachloro-1, 1', 3, 3'-tetraethylbenzimidazolylcarbocyanine iodide+%, JC-1+%) was used to indicate the change of normal MMP.Results The death rate of BRL hepatocytes increased significantly after 72 h induced by 400 μmol/L PA. PA decreased the expression of SCD1 in CON+ group and NC+ group, but it increased the expression in SCD1-LV+group and SCD1-LV group. JC-1+% was lower in CON+ and NC+ groups, but JC-1+% in SCD1-LV+ group was similar to that in control group.Conclusions The expression of SCD1 increases in BRL hepatocytes when they are infected by SCD1 overexpression lentiviral vector. The destaturation of saturated fatty acid is enhanced. Overexpression of SCD1 decreases the toxicity induced by PA. The MMP disorder of infected BRL hepatocytes is retrieved.
Keyword:
Stearoyl-CoA desaturase-1; Palmitic acid; Lentiviral vector; Mitochondrium membrane potential
饱和脂肪酸过多的摄入会引起多种代谢性疾病, 单不饱和脂肪酸具有重要的生理功能。硬脂酰辅酶A去饱和酶1(stearoyl-CoA desaturase-1, SCD1)主要催化棕榈酰辅酶A和硬脂酰辅酶A去饱和为棕榈油酰辅酶A和油酰辅酶A, 从而减少饱和脂肪酸的量, 降低脂毒性。动物实验中我们观察到随着高脂喂养时间的延长, SD大鼠肝细胞SCD1表达下调及细胞凋亡增加, 线粒体膜电位(mitochondrium membrane potential, MMP)紊乱是细胞凋亡的重要特征。结合其他文献我们推测上调SCD1表达可能对缓解高脂引起的肝细胞凋亡有帮助, 为此我们设计如下实验。通过大鼠BRL肝细胞系和棕榈酸(PA)共培养, 以观察BRL肝细胞系SCD1表达变化、细胞凋亡及MMP的改变。然后通过已经构建成功的pGC-FU-SCD1-GFP慢病毒载体转染大鼠BRL肝细胞系, 检测在SCD1过表达状态下, 是否能够降低细胞内饱和脂肪酸引起的脂毒性和恢复MMP, 从而减少细胞的凋亡和膜电位的下降, 进一步阐明SCD1在肝细胞能量调节中的作用及其机制, 为临床非酒精性脂肪肝的治疗提供帮助。
材料和方法
一、材料
1. 大鼠BRL肝细胞系 大鼠BRL肝细胞系购于上海中科院细胞所。
2. SCD1过表达慢病毒载体 pGC-FU-SCD1-GFP慢病毒载体已构建成功[1]。
3. 主要试剂 活体细胞膜功能(膜电位)荧光测定试剂盒购自上海杰美基因科技有限公司, 台盼兰购自上海捷倍思基因技术公司, PA、低脂肪酸的牛血清白蛋白V(BSA)购自美国Sigma公司, 引物合成于上海生工生物工程有限公司。
4. 仪器 FACS Calibur流式细胞仪为美国Becton Dickinson公司产品, 实时聚合酶链反应(PCR)仪为美国Stratagene产品。
二、方法
1. PA溶液的配制[2] PA溶液为脂溶性, 必须有BSA助溶。称取372 mg PA溶解在5 mL无水乙醇中, 震荡5 min后完全溶解, 再加入5 mL双蒸水配制成母液, 将母液放进50 ℃恒温水浴箱孵育10 min。取母液1.334 mL溶解在含10% BSA的RPMI-1640培养基中, 至10 mL, 在37 ℃水浴箱搅拌, 待完全溶解, 超净台下微量滤器除菌, 然后溶解于改良Eagle培养基(DMEM)中, 配制0.2、0.4、0.8 mmol/L的PA孵育液, 并置4 ℃保存。
2.高脂诱导下BRL肝细胞死亡率检测 取24孔板接种细胞, 每组设9个复孔, 于培养箱中培养。当细胞融合率为50%时分组。以无PA培养的BRL细胞作为对照, 分别用200、400和800 μ mol/L的PA孵育液共培养72 h。胰酶消化, 0.4%台盼兰染色2~3 min。取细胞悬浮液滴加到细胞计数板, 静置15 min, 镜下观察, 计细胞死亡率。取细胞死亡率下降显著对应的PA浓度进行下游试验。
3.MMP的检测 实验分为加药组和非加药组。非加药组包括一般培养组(CON)、一般培养加阴性病毒组(NC)和一般培养加过表达病毒组(SCD1-LV); 加药组包括一般培养加PA组(CON+)、一般培养加阴性病毒和PA组(NC+)、一般培养加过表达病毒和PA组(SCD1-LV+)。每组接种1× 105个BRL细胞, 每组3个复孔, 实验重复3次。当细胞融合率达到30%时按照预实验复感染指数(MOI)=20加入适量的病毒液。72 h后收集细胞于15 mL尖底离心管; 加入500 μ L胰酶, 铺满整个培养孔, 消化移入15 mL锥形离心管, 2 590 r/min(离心半径为4 cm)离心5 min, 抽去上清液。加入50 μ L 清理液(reagent E), 混匀细胞颗粒群, 移入1.5 mL EP管中。加入450 μ L 含有GENMED染色液B(reagent C)和GENMED稀释液(reagent D)的染色工作液, 轻度涡旋震动几秒钟。37 ℃ 细胞培养箱孵育20 min(注意避光操作); 2 590 r/min(离心半径为4 cm)离心5 min, 抽去上清液。加入500 μ L清理液(注意检查前置于冰槽), 混匀后即刻进行流式细胞仪单通道分析。采用藻红蛋白(phycoerythrin, PE)通道, 激发波长488 nm, 散发波长575 nm。用发橙红色荧光细胞百分率(JC-1+%)表示MMP正常细胞比例。正常细胞有一定的红色荧光强度, 膜电位受到损伤时, 强度则显著减弱。
4. SCD1 mRNA检测 各组细胞模型建立方法同“ 方法3” 。细胞总RNA抽提和逆转录, 实时PCR检测SCD1表达, 引物序列为SCD1 上游引物:5'-AAAGAGAAGGGCGGAAAGC-3', 下游引物:5'-GTGTGGGCAGGATGAAGC-3'(115 bp); 3-磷酸甘油醛脱氢酶(glyceraldehyde 3 phosphate dehydrogenase, GAPDH)上游引物:5'-TTCAA-CGGCACAGTCAAGG-3', 下游引物:5'-CTCAGCA-CCAGCATCACC-3'(114 bp)。反应条件:预变性95 ℃ 15 s、变性95 ℃ 5 s、退火延伸60 ℃ 30 s, 45个循环; 每次在延伸阶段读取吸光度值。
三、 统计学方法
采用SPSS 13.0软件进行分析, 所有数据以
结 果
一、 BRL肝细胞在PA诱导下的存活率测定
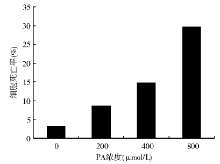
随着PA浓度的增大和培养时间的延长, 细胞存活率有明显下降。与对照组相比, 400 μ mol/L PA处理72 h诱导了14.8%的细胞死亡(P< 0.01); 800 μ mol/L PA处理72 h诱导了29.6%的细胞死亡(P< 0.01)。见图1。
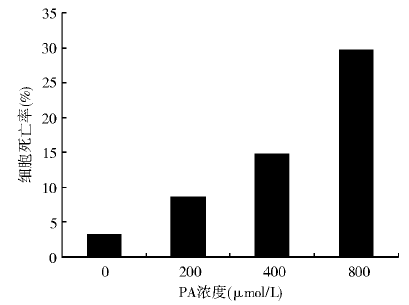 | 图1 不同浓度PA处理72 h对BRL肝细胞死亡率的影响 |
二、 各组BRL肝细胞系SCD1 mRNA表达
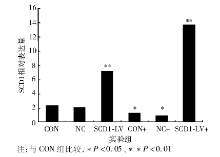
实时PCR结果显示, 与CON和NC组相比, CON+和NC+组SCD1 mRNA表达明显下降, SCD1-LV和SCD1-LV+表达上升, SCD1-LV+表达升高尤为明显。见表1和图2。
| 表1 各组BRL肝细胞SCD1表达分析结果 ( |
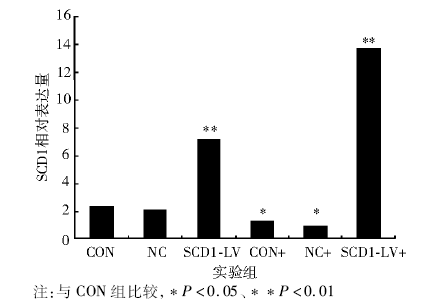 | 图2 各组BRL肝细胞SCD1的表达变化 |
三、各组BRL肝细胞系MMP检测结果
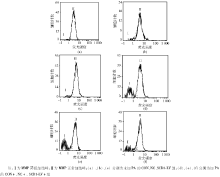
流式细胞分析仪结果显示不加PA的CON、NC和SCD1-LV组MMP正常细胞比例分别为97.9%、96.2%和96.8%; 加PA的CON+、NC+和SCD1-LV+组MMP正常细胞比例分别为76.8%、73.2%和88.5%。CON+、NC+组MMP正常细胞的比例明显下降, SCD1-LV+组JC-1+%高于CON+、NC+组(P< 0.05)。见图3。
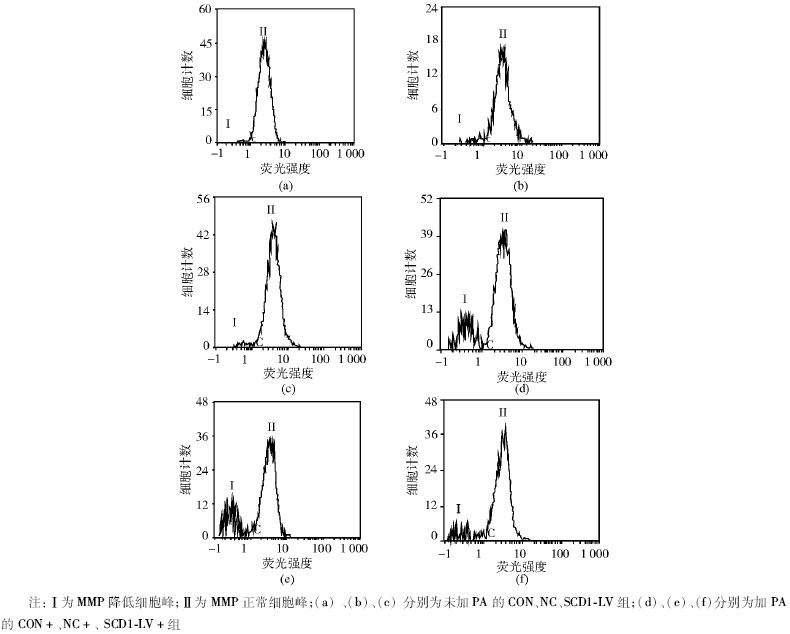 | 图3 各组BRL肝细胞MMP检测结果 |
讨 论
脂质代谢异常是一种重要的代谢综合征, 涉及了非酒精性脂肪肝、肝硬化、心血管疾病、糖尿病和肥胖等多种疾病危险因素, 已经成为医学上研究的热点。脂毒性是指血中游离脂肪酸(free fatty acid, FFA)超过脂肪组织储存能力和各组织对FFA的氧化能力, 使过多的FFA以三酸甘油酯形式在非脂肪组织中过度沉积而造成对该组织的损伤。肝脏是脂代谢的最重要器官。FFA参与脂肪肝形成的同时造成了肝细胞凋亡损伤乃至坏死, 从而导致肝脏功能障碍。FFA包括饱和脂肪酸和不饱和脂肪酸。FFA除了作为机体功能的重要物质外, 其在细胞膜生物合成和细胞内信号转导过程中也具有重要的作用, 是重要的脂质毒性物质。饱和脂肪酸诱导的脂凋亡在多种细胞中也得到证实, 如中国仓鼠卵巢细胞[3]、心肌细胞[4]、乳腺癌细胞[5]及胰岛β 细胞[4]等。关于脂凋亡的机制存在多种学说, 但研究普遍认为FFA的脂凋亡作用与其饱和度有关。饱和脂肪酸如PA诱导β 细胞脂性凋亡, 而不饱和脂肪酸如油酸则较少细胞毒性, 并且能够保护饱和脂肪酸诱导的凋亡作用[6]。
本研究采用血浆中含量丰富的PA作为试验的干预因素诱导大鼠BRL肝细胞系凋亡, 用SCD1重组(pGC-FU-SCD1-GFP)慢病毒载体诱导SCD1的过表达, 探讨SCD1表达降低饱和脂肪酸引起细胞凋亡的机制。先用慢病毒感染72 h后加PA诱导。PA诱导72 h后, SCD1表达下降, 可能与高脂诱导后引起内源性高瘦素表达水平有关[7]; SCD1基因是瘦素信号的靶基因, 瘦素通过转录因子甾醇调节单元结合蛋白(sterol regulatory element binding protein, SREBP)[8]或神经肽(neuropeptide Y, NPY)[9]发生作用, 抑制SCD1基因的表达; 而SCD1-LV和SCD1-LV+组与CON及NC组相比, SCD1 mRNA表达明显上升(P< 0.01); 并且SCD1-LV+组比SCD1-LV组表达更为显著, 可能是由于PA刺激增强SCD1的表达。
MMP的形成主要与质子泵有关, 经三羧酸循环产生的还原型烟酰胺腺嘌呤二核苷酸(NADH)和还原型黄素腺嘌呤二核苷酸(FADH2)在氧化过程中, 其高能电子在呼吸链上传递, 释放的能量使质子(H+)通过线粒体内膜上的质子泵从线粒体内膜的基质侧移至膜间腔。线粒体内膜对H+不能自由通透, 从而在线粒体内膜两侧形成电化学质子梯度, 即膜电位, 通常反映线粒体膜通道(PTP孔)开放的情况。凋亡过程中, PTP孔的开放可使线粒体内膜通透性突然增加, 这种通透性可导致MMP的崩解。PTP孔的开放还可以使线粒体腔内的细胞色素C释放到细胞浆中, 细胞色素C与凋亡酶激活因子-1 (apoptotic protease-activating factor 1, Apaf-1)结合, 在脱氧腺苷三磷酸/腺苷三磷酸(ATP)存在时, 该复合物结合并激活半胱天冬酶9酶原, 再激活下游的半胱天冬酶(如半胱天冬酶3等)进而引发凋亡。线粒体是神经酰胺介导凋亡的靶器官[10, 11], 线粒体膜屏障受损的半胱天冬酶途径也是一个重要因素。Maedler等[12]发现PA可降低成年鼠β 细胞线粒体腺嘌呤核苷酸转位分子(ANT)的表达, ANT可催化ATP和腺苷二磷酸(ADP)通过内膜交换, ANT下降, 引起ATP和ADP交换受阻, 线粒体肿胀, 外膜受损, 胞内细胞色素C进入胞质, 激活半胱天冬酶途径, 引起细胞凋亡及MMP的紊乱。对培养的BRL肝细胞用流式细胞分析仪分析, 发现未加入PA的CON、NC、SCD1-LV组JC-1+%维持在较高的水平, 而PA刺激的CON+和NC+组JC-1+%显著下降, SCD1过表达PA刺激的SCD1-LV+组 JC-1+%明显高于CON+、NC+组, 但与CON组无明显差异。此结果表明, SCD1的过表达能够有效的催化PA为单不饱和脂肪酸, 使MMP的紊乱得到恢复, 从而降低脂毒性。
本研究以PA为干预因素, 诱导大鼠BRL肝细胞脂凋亡, 然后利用SCD1慢病毒载体过表达SCD1, 发现SCD1的过表达能够降低PA脂毒性, 纠正饱和脂肪酸引起的MMP下降, 说明SCD1可能通过维持线粒体膜的稳定降低饱和脂肪酸诱导的BRL肝细胞的凋亡, 为临床上治疗肝脏脂肪病变甚至代谢综合征提供了新的思路 。
The authors have declared that no competing interests exist.
参考文献
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|